View in English?
View in English?
View in English?
正常な免疫システムでは、自己免疫寛容を維持し免疫応答に伴う組織の損傷を防ぐため、一連の免疫チェックポイント機構が備わっています。CD8+細胞障害性T細胞や、CD4+ヘルパーT細胞の活性化には、T細胞受容体 (TCR) と抗原の相互作用が必要で、抗原ペプチドは抗原提示細胞 (APC) 表面に、主要組織適合複合体 (MHC) に結合した形で提示されます (1)。さらに、T細胞上の共刺激分子 (CD28、ICOS、GITRなど) と、抗原提示細胞上の受容体 (CD80/CD86、ICOSL、GITRLなど) の相互作用も、T細胞の活性化に関与しています (1)。しかし、特定の環境下では、T細胞受容体や共刺激分子の会合は、T細胞の活性化と免疫応答を抑制する、阻害シグナルにもなります。このような阻害シグナルには、PD-1、CTLA-4、TIM-3、LAG3 (2) といった、免疫チェックポイント制御タンパク質が関与しています。免疫チェックポイントタンパク質PD-1とCTLA-4は、腫瘍浸潤T細胞で共通して増加しており、対応するリガンドであるPD-L1 (B7-H1)/PD-L2 (B7-DC) とCD80/86にそれぞれ結合してT細胞応答を抑制します。免疫チェックポイントのリガンドはがん細胞でしばしば高発現が見られ、がん細胞はこれによって免疫を回避していると考えることができます (3-5)。このため、免疫チェックポイントを阻害して抗腫瘍免疫を活性化することが、創薬やがん治療法の開発の新たなアプローチとして注目されています (例:抗PD-1免疫療法) (6)。
下の表は、T細胞応答の活性化と抑制に関与する、促進性および抑制性の受容体-リガンド複合体の一覧です。詳細情報や関連製品一覧については、リンクのあるタンパク質名をクリックしてください。
| 細胞応答 | T細胞 | 抗原提示細胞 |
|---|---|---|
| 共刺激性 | CD28 | B7-1 (CD80) or B7-2 (CD86) |
| CD40L | CD40 | |
| TLT-2?** | B7-H3 | |
| OX40 (CD134) | OX40L | |
| 4-1BB (CD137) | 4-1BBL | |
| ICOS | ICOSL | |
| GITR | GITRL | |
| 共抑制性 | CTLA-4 | B7-1 (CD80) or B7-2 (CD86) |
| PD-1 | B7-H1 (PD-L1) or B7-DC (PD-L2) | |
| 不明 | B7-H3 | |
| 不明 | B7-H4 | |
| 不明 | VISTA | |
| VISTA | 不明 | |
| LAG3 | MHC-Class II | |
| TIM-3 | Galectin-9 |
**Myeloid cell-like transcript2 (TLT-2) は、B7-H3 (7,8) の推定上の受容体とされています。ただし、その共刺激の意義や、それが実際にB7-H3と相互作用するかどうかについては疑問視されてきています (9)。従って、この受容体とリガンドの相互作用の妥当性は議論を呼ぶところとなっています。
また、IDOやArginase-1といった免疫代謝関連タンパク質の役割にも関心が高まりつつあり、創薬ターゲットの候補分子として期待されています。IDOは、多くの腫瘍で増加が見られるトリプトファン分解に関わる免疫抑制性の酵素です。Arginase-1は、遍在する酵素ですが、腫瘍微小環境へとリクルートされる骨髄由来免疫抑制細胞 (MDSC) でしばしば高発現しています。Arginase-1は、L-arginineの尿素への変換を触媒し、それによりT細胞の活性化や分化に必須の栄養素を奪います (10)。
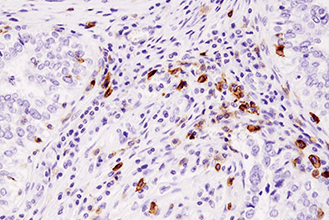
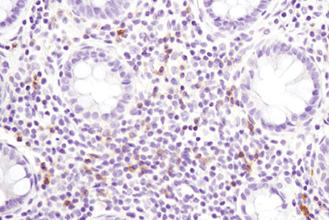
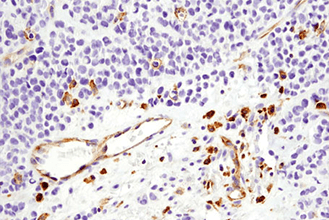
がんのような、生理学的にも病理学的にも複雑な条件下にある組織の微小環境を理解するためには、免疫チェックポイントタンパク質および表現型マーカーをプロファイリングすることが重要です。この際、限られた貴重な患者サンプルで、複数のがん免疫療法の標的分子やバイオマーカーを同時に調べられるマルチプレックスアッセイが有効です。下の表は、免疫細胞表現型マーカーの一覧です。詳細情報や関連製品一覧については、リンクのあるタンパク質名をクリックしてください。
| 表現型&機能マーカー | |
|---|---|
| CD3ε | CD4 |
| CD5 | CD8α |
| CD11c | CD14 |
| CD16 | CD19 |
| CD31 | CD36 |
| CD44 | CD45 |
| CD45RO | CD56 (NCAM) |
| CD68 | CD79A |
| CD79B | CD163 |
| CD206 | T-BET |
| F4/80 | FOXP3 |
| Arginase-1 | IDO |
| GATA-3 | Granzyme B |
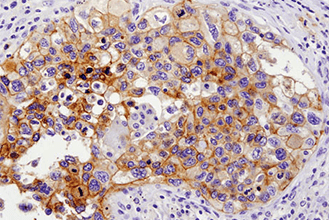
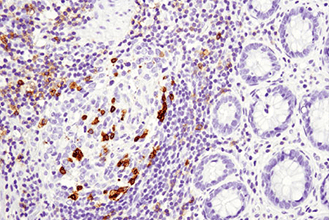
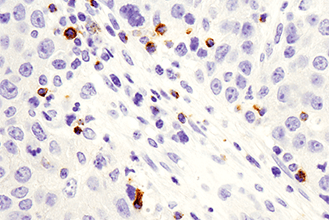
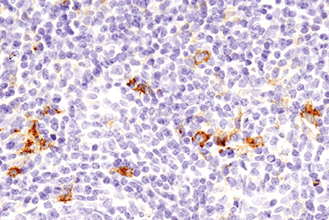
複数のタンパク質やバイオマーカーの検出が要求される、腫瘍免疫学的アプローチには、ホルマリン固定パラフィン包埋 (FFPE) 組織サンプルで6種類以上の標的を同時検出可能な、蛍光マルチプレックス免疫組織化学染色 (mIHC) が有効です。mIHCにおいて信頼度の高い結果を得るためには、単色/二色の発色法を用いたIHCアプローチと同様に、適切なターゲットに対する検証済みの抗体を用いることが極めて重要です。CSTは、ヒトおよびマウスへ交差するIHC検証済み抗体をご提供しており、これらを用いることで、バイオマーカーの発現、局在、相互作用、病態に関する詳細な情報を得ることができます。
View in English?