View in English?
View in English?
View in English?
CSTのシグナル伝達パスウェイ図で個々のタンパク質名をクリックすると、それに関連した研究リソースや製品情報を検索することができます。さらに、教育・研究のために、パスウェイ図をダウンロードすることも可能です。

受胎の直後、受精卵 (接合子) は胚発生、すなわち高度に制御された増殖と方向づけられた分化を開始します。胚発生時の様々なイベントは、複雑な遺伝的、およびエピジェネティックなシグナル伝達カスケードによって導かれます。それは、完全な多細胞生物を一から形作る非常に複雑なプロセスの第一歩です。
胚性幹細胞 (ESC) は、卵割期胚 (胚盤胞) の内部細胞塊より誘導される細胞集団として定義されます。ESCの決定的な特徴は、それらに固有の多分化能です。身体のどんな細胞系統へも分化することができ、無限の自己複製が可能になります。ESCのこれらの特徴は、複雑な一連の細胞シグナル伝達カスケードにより緊密に制御されています。発生生物学における強力な研究ツールとして活用され、さらに、個別化再生医療における潜在性も見出されています。
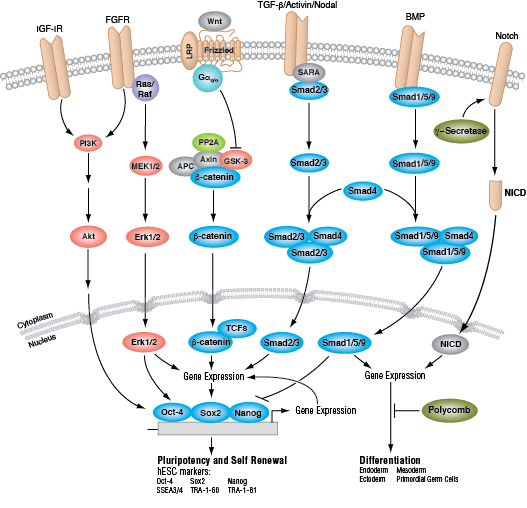
ヒトでは、ESCにおける多分化能および自己複製能の維持を担う主要なシグナル伝達経路が2つ知られており、それは、SMADタンパク質を介したBMP/TGF-βシグナル伝達経路と、MAPKとAktの経路を活性化するFGFシグナル伝達経路です。Wntシグナル伝達経路も多分化能を促進しますが、これは転写活性化因子であるTCF1と、転写抑制因子であるTCF3との間の均衡に関与する非古典的メカニズムを介して行われている可能性があります。これらの経路を介するシグナル伝達は、3つの主要な転写因子の発現および活性化をもたらします。それは、Oct-4、Sox2、NANOGの3つです。これらの転写因子は、ESC特異的遺伝子の発現を促し、また自らの発現も制御し、さらに多分化能の有用なマーカーとしても使用されます。hESCの同定に使われるその他のマーカーは、細胞表面の糖脂質であるSSEA3/4、ならびに糖タンパク質のTRA-1-60およびTRA-1-81を含みます。
人工多能性幹細胞 (iPSC) は、多分化能を有するESC様の細胞であり、所定の「再プログラミング」因子セットの強制発現により、分化した細胞から誘導することができます。そのセットで最もよく知られているのはOCT-4、SOX2、KLF4、c-MYCです。再プログラミングが成功すると、iPSCはESCと同様の遺伝子発現シグネチャーを示し、多分化能および自己複製能の両方を示します。これにより、iPSは研究コミュニティからの大きな注目を集めてきています。なぜならば、研究にiPSを使用することができれば、ヒト胚盤胞から誘導されたESCの使用を取り巻く倫理的および技術的な問題点の多くを回避することができるからです。ESCと同様に、iPSCは、再生医療・個別化医療、薬物スクリーニングでの利用において潜在性があり、さらに、胚発生を制御する細胞シグナル伝達カスケードの理解を更に深められることが期待されて、現在、中心的な研究対象になっています。
ESCおよびiPSCのいずれにおいても、原腸形成の間に確立される3つの一次胚葉の細胞タイプ(外胚葉、中胚葉、および内胚葉) への成長を誘導することができます。外胚葉は神経幹細胞の前駆体であり、細胞分裂して脳、脊髄、および末梢神経を含む細胞を産生します。外胚葉起源の他の細胞は、表皮、および消化管の末端領域を含みます。中胚葉は、脂肪、筋肉および骨、造血幹細胞の前駆体である、間葉系幹細胞に分化します。造血幹細胞は、血液および免疫システムの全ての細胞系統を産生します。内胚葉は、内胚葉前駆細胞 (肝と膵細胞の両方の前駆体) に分化します。これは、消化管、気道および尿路を覆う細胞の前駆体でもあります。
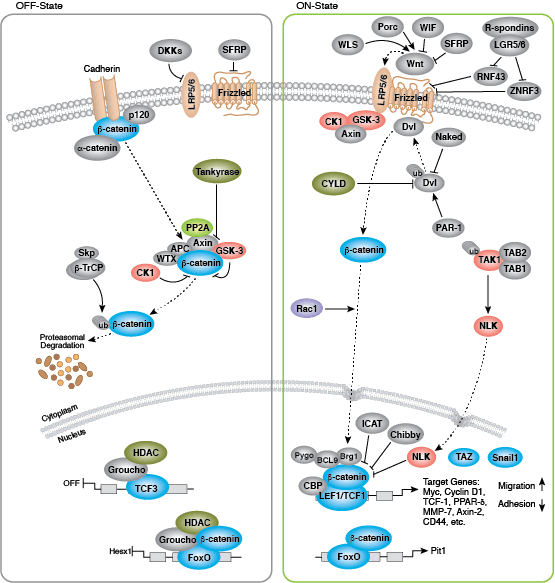
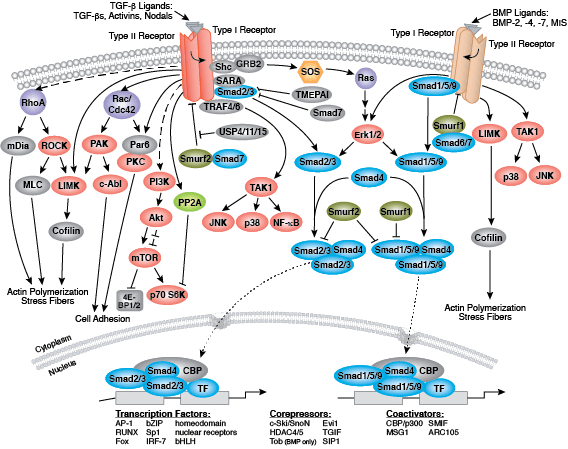
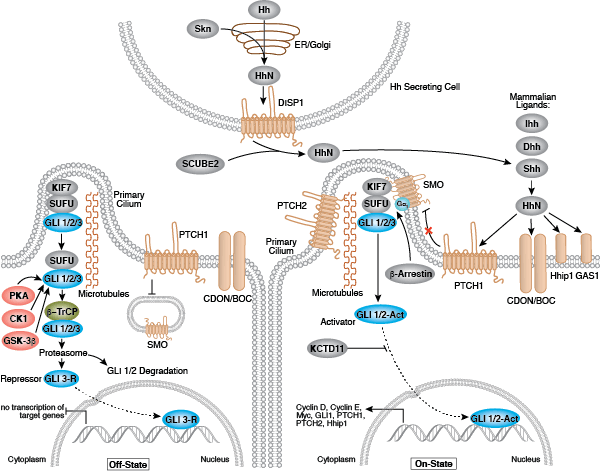
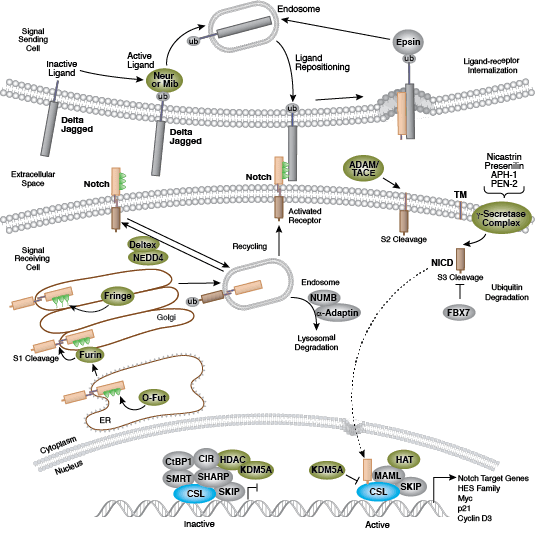
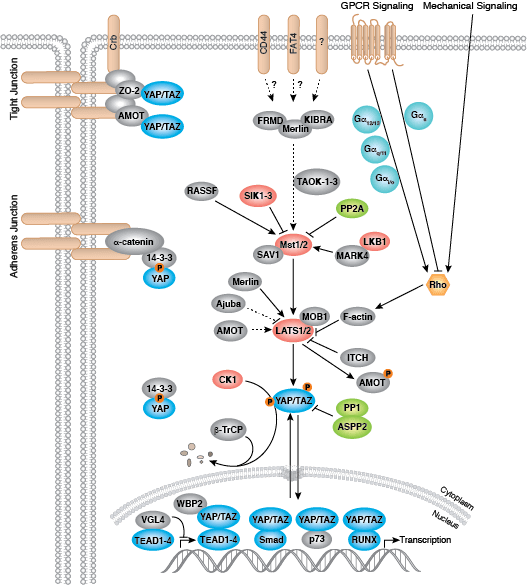
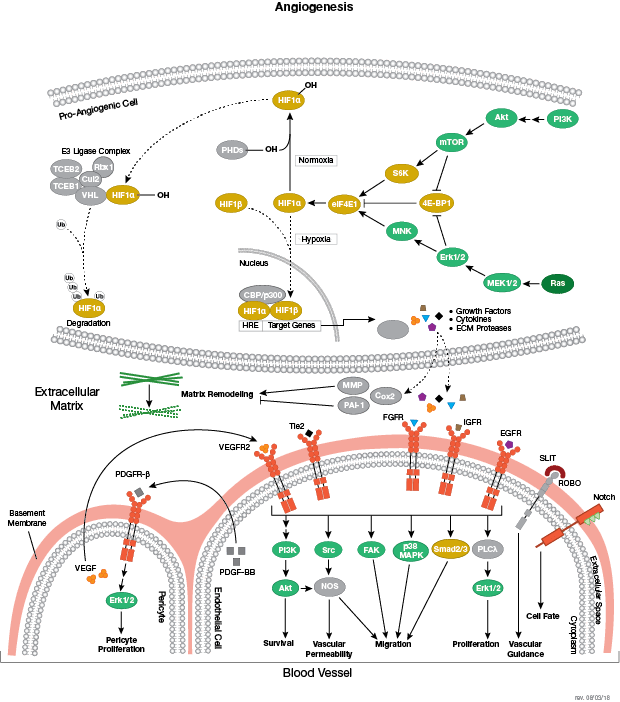
それぞれの分化系統への発生は、BMP/TGF-β、Notch、Wnt/β-catenin、Headgehog、Hippo経路を含む、細胞分裂、増殖および分化を制御するいくつかのシグナル伝達経路により制御されます。これらの経路はそれぞれ、複雑な一連の遺伝的、もしくはエピジェネティックな (例えば、ヒストンの修飾)、あるいは外因性のシグナル伝達因子により制御され、発生・分化の間の細胞運命や細胞の振る舞いを導きます。
View in English?