View in English?
View in English?
View in English?
DNAメチル化やヒストン修飾といったエピジェネティック制御の異常は、アルツハイマー病、ハンチントン病、パーキンソン病、多発性硬化症、筋萎縮性側索硬化症に関与しています。しかしながら、疾患の進行に対するこれらの詳細な影響は未だ明らかにされていません。脳の健康状態はエピジェネティックなメカニズムに大きく依存しており、神経変性疾患ではクロマチン動態に障害が見られます。環境の改善とエピジェネティックスの変化による発症リスクに着目した研究は、神経変性疾患の治療法の開発にて成長している分野です。
5-ヒドロキシメチルシトシン (5-hmC) は、Tetタンパク質ファミリーによる5-mC (5-メチルシトシン) の酸化によって生じる新たな修飾シトシンです。発生を通じた5-hmCのパターンは、適切な神経発生や神経機能に不可欠です。この調節不全は、アルツハイマー病やハンチントン病などの神経変性疾患を引き起こしますが、その正確なメカニズムは未だ明らかにされていません。
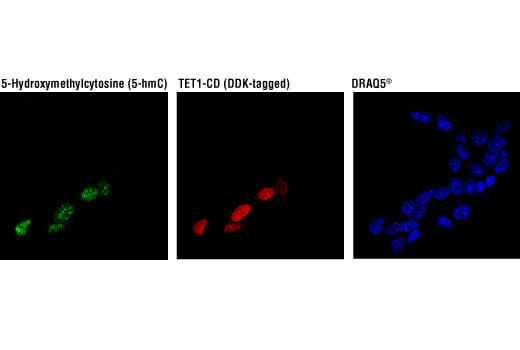
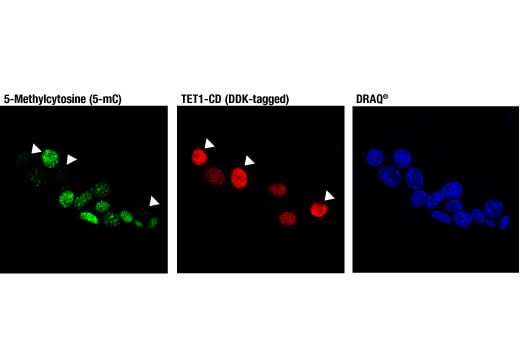
5-メチルシトシン (5-mC) は、未修飾状態以降の脳のシトシンの最も一般的な状態です。細胞の加齢に伴い、脳のゲノム全体での5-mC含有量は減少します。アルツハイマー病のニューロンではゲノム全体に5-mCの減少が見られ、筋萎縮性側索硬化症やパーキンソン病の進行にも5-mCの調節不全が関与している可能性があります。
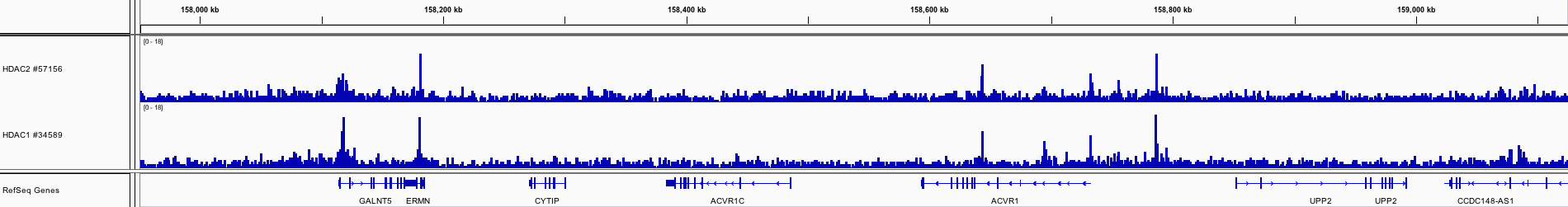
HDAC2はクラスIヒストン脱アセチル化酵素であり、通常、遺伝子発現を抑制します。HDAC2を欠損したアルツハイマー病のマウスモデルでは、認知機能の向上とアミロイドの蓄積の低減が見られます。また、アルツハイマー病患者ではHDAC2の発現増加が見られます。このほか、HDAC2はハンチントン病や多発性硬化症にも関与しています。
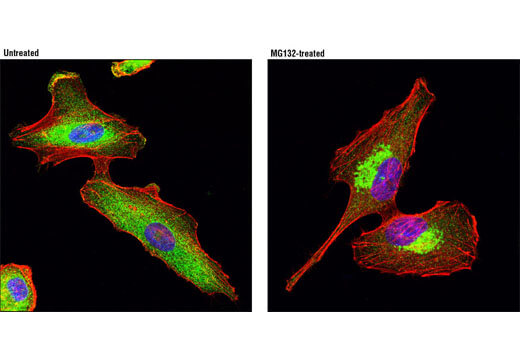
HDAC6はクラスIIヒストン脱アセチル化酵素で、アルツハイマー病患者の大脳皮質や海馬で発現の増加が見られます。HDAC6はTauタンパク質と共局在し、Tauのリン酸化と相関します。HDAC6の減少が認知機能の向上につながる可能性があります。
p300はヒストンアセチル化酵素で、神経活性に応答して制御されるクロマチンのアセチル化に重要な役割を果たします。アルツハイマー病のマウスモデルのニューロンではヒストンアセチル化レベルの低下が見られ、ニューロンの初代培養系ではアミロイド前駆体タンパク質に依存的なシグナル伝達の活性化によってヒストンアセチル化酵素の減少が見られます。p300はハンチントン病やパーキンソン病にも関与しています。
View in English?