

クロマチン免疫沈降 (ChIP) アッセイの方法論概要
ChIPアッセイの概要
クロマチン免疫沈降 (ChIP) アッセイは、細胞に自然な状態で存在するクロマチンのタンパク質とDNAとの相互作用を解析するための、強力かつ多機能な手法です (1,2)。このアッセイは、特定のゲノム領域に結合した複数のタンパク質の同定、またはその逆に、特定タンパク質が結合する複数のゲノム領域を同定するために使用することができます (3–6)。さらに、ChIPアッセイでは、特定のタンパク質-DNA相互作用の、空間的・時間的な関連性を解析することができます。例えば、ある遺伝子のプロモーター領域に結合するタンパク質因子群が、どのような順番に結合していくのかを特定する場合にChIPアッセイを用いることができます。また、ある遺伝子が活性化されるときに、遺伝子座全体にまたがる特定のヒストン修飾の相対量を測定する場合に用いることもきます (3,4)。さらに、、ChIPアッセイはヒストンタンパク質のほか、転写因子、転写コファクター、DNA複製因子、DNA修復タンパク質の結合の解析に用いることができます。
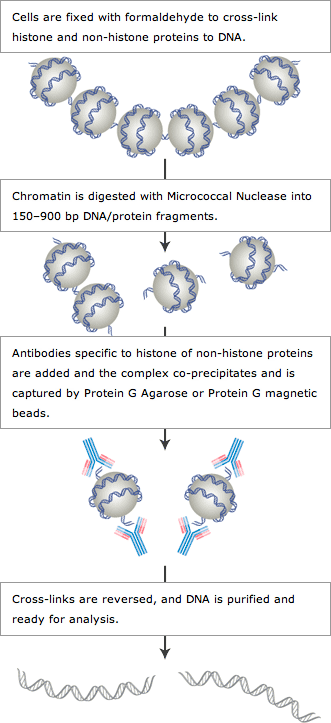
ChIPアッセイでは、はじめに細胞をホルムアルデヒド (可逆的なタンパク質ーDNAクロスリンク剤) で処理します。これによって細胞内で起こっているタンパク質ーDNAの結合が固定され、実験操作中も両者の結合が保持されます (方法の概要をご参照ください) (1,2)。その後、細胞を溶解して回収したクロマチンを、ソニケーションあるいは酵素消化によって断片化します。さらに、特定のタンパク質またはヒストン修飾に対して特異的な抗体を用いて、クロマチンを免疫沈降します。目的のタンパク質と結合するDNA配列や、目的のヒストン修飾を受けたDNA配列は、クロスリンクされたクロマチン複合体の一部として共沈降します。すなわち、免疫沈降という選択的なプロセスを経ることで、目的のDNA配列の相対量が濃縮されます。免疫沈降後、タンパク質とDNAを脱クロスリンクし、DNAを精製します。濃縮されたDNA配列は、目的に応じて、次のような方法で検出します。
目的のタンパク質が結合したDNA配列や領域、目的のヒストン修飾を受けたDNA配列や領域を同定するのによく用いられる手法として、標準的なPCR法が挙げられます (1,2)。PCR法では、目的のタンパク質に特異的な抗体を用いた免疫沈降で濃縮された特定のDNA配列の相対量を測定し、非特異的なコントロール抗体を用いた場合と比較します。PCR産物を、アガロースゲルまたはアクリルアミドゲルで電気泳動して定量し、特定のDNA配列の濃縮レベルをinput DNAの総量に対して算出します (percent of input)。濃縮レベルは、非特異的抗体コントロールでみられる濃縮 (バックグラウンド) に対して何倍濃縮されるか、としても表すこともできます。リアルタイムPCRは、ゲルによる電気泳動を行わずに、特定のDNAの濃縮レベルをより正確に定量するシステムです。また、ChIPアッセイは、ゲノムタイリングマイクロアレイ (チップオンチップ) 法、NGSによるシーケンシング、あるいはクローニング法と組み合わせることで、タンパク質-DNA相互作用やヒストン修飾を、ゲノムワイドで解析することが可能になります (5–8)。
CSTの細胞サンプル用、組織サンプル用酵素法ChIPキットには、哺乳動物細胞を用いたChIPアッセイに必要なバッファー類、試薬類が含まれており、これはヒストン修飾の解析と非ヒストンDNA結合タンパク質の解析のどちらでも使用できます。細胞溶解後、クロマチンをMicrococcal Nucleaseを用いた部分消化によって断片化し、ヌクレオソーム1つから5つ分に相当するサイズのクロマチン断片を調製します。酵素消化によるクロマチンの断片化は、ソニケーションによる断片化よりはるかに穏やかな手法です。また、ソニケーション中のクロマチンの乳化の問題を回避する、ソニケーション強度のバラツキによる断片化レベルのバラツキの問題を回避するなどの利点があります。これらの問題は、不完全なクロマチン断片化、またはタンパク質の変性や分解による抗体エピトープの損失の原因となる可能性があります。クロマチンの断片化の後、ChIP Grade Protein G AgaroseまたはChIP Grade Protein G Magnetic Beadsのいずれかと、目的の抗体を用いて、クロマチン免疫沈降を行います。免疫沈降後、タンパク質とDNAの脱クロスリンクを行い、キットに含まれるDNA精製スピンカラムを用いてDNAを精製します。DNA精製のステップは、独自に設計されたシリカ膜の特異的な結合を利用して、高効率かつ簡便に行うことができ、タンパク質の夾雑物を除去に、フェノール/クロロホルム抽出やエタノール沈殿を必要としません。DNA精製後、特定のDNA配列の濃縮は、標準的なPCR法や定量的リアルタイムPCRで特定のDNA配列の濃縮レベルを解析することができ、またチップオンチップ、NGSによるシーケンシングやクローニング技術などの解析に用いることも可能です。
方法の概要
SimpleChIP®キットには、ChIPアッセイに必要なバッファーと試薬だけでなく、ChIP実験が成功しているかの判断を可能にする重要なコントロールが含まれています。このキットにはポジティブコントロールのHistone H3抗体、ネガティブコントロールのNormal Rabbit IgG、そしてRibosomal protein L30 (RPL30) 遺伝子座をPCRで検出するプライマーセット (ヒトおよびマウスのプライマーセット) が含まれています。Histone H3は、細胞でクロマチンの主要な構成因子で、RPL30遺伝子座を含むゲノム全体のほとんどのDNA配列に結合します。つまり、Histone H3抗体を用いてクロマチン免疫沈降を行なった場合はRPL30遺伝子の濃縮が起こる一方、Normal Rabbit IgGを用いた場合には濃縮が起こりません。キットに付属するRPL30プライマーセットを使用して、標準的PCRまたは定量的リアルタイムPCRを行うことで、RPL30遺伝子の濃縮レベルを評価することができます。重要なことは、Histone H3はゲノム全体のほとんどのDNA座にも結合するため、Histone H3抗体はおよそどんな遺伝子座の研究のためのポジティブコントロールのIPとして利用可能で、ユーザーはChIP実験の成功を確認することができます。
SimpleChIP®キットは、最大6回までのクロマチン調製 (または最適化) および 30免疫沈降に十分な試薬を含み、4X 107細胞を用いたスケールの実験系に最適化されています。ChIPアッセイは2日間程度で実施可能で、使用する細胞数に合わせて、スケールアップ/ダウンが簡単にできます。
参考文献
- Orlando V (2000) Mapping chromosomal proteins in vivo by formaldehyde-crosslinked-chromatin immunoprecipitation. Trends Biochem. Sci. 25(3), 99–104.
- Kuo MH, Allis CD (1999) In vivo cross-linking and immunoprecipitation for studying dynamic Protein:DNA associations in a chromatin environment. Methods 19(3), 425–33.
- Agalioti T, Lomvardas S, Parekh B, Yie J, Maniatis T, Thanos D (2000) Ordered recruitment of chromatin modifying and general transcription factors to the IFN-beta promoter. Cell 103(4), 667–78.
- Soutoglou E, Talianidis I (2002) Coordination of PIC assembly and chromatin remodeling during differentiation-induced gene activation. Science 295(5561), 1901–4.
- Mikkelsen TS, Ku M, Jaffe DB, Issac B, Lieberman E, Giannoukos G, Alvarez P, Brockman W, Kim TK, Koche RP, Lee W, Mendenhall E, O'Donovan A, Presser A, Russ C, Xie X, Meissner A, Wernig M, Jaenisch R, Nusbaum C, Lander ES, Bernstein BE (2007) Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 448(7153), 553–60.
- Lee TI, Jenner RG, Boyer LA, Guenther MG, Levine SS, Kumar RM, Chevalier B, Johnstone SE, Cole MF, Isono K, Koseki H, Fuchikami T, Abe K, Murray HL, Zucker JP, Yuan B, Bell GW, Herbolsheimer E, Hannett NM, Sun K, Odom DT, Otte AP, Volkert TL, Bartel DP, Melton DA, Gifford DK, Jaenisch R, Young RA (2006) Control of developmental regulators by Polycomb in human embryonic stem cells. Cell 125(2), 301–13.
- Weinmann AS, Farnham PJ (2002) Identification of unknown target genes of human transcription factors using chromatin immunoprecipitation. Methods 26(1), 37–47.
- Wells J, Farnham PJ (2002) Characterizing transcription factor binding sites using formaldehyde crosslinking and immunoprecipitation. Methods 26(1), 48–56.