

代謝と代謝疾患の概要
代謝と代謝異常
代謝 (Metabolism) という言葉は「変化すること」を意味するギリシャ語を起源としており、恒常性を維持する細胞、組織、器官で起こる化学変化またはプロセス全般を指します。ここでは、細胞レベルの異なる代謝経路と、これらに障害が起きた場合の代謝疾患の病態形成にフォーカスして総説します。代謝変化の根底を成す様々なシグナル伝達イベントや化学反応を解析することで、正常細胞の生理学と疾患の理解を深めることができます。ヒトの疾患で起こる代謝の異常がどのように起こっているのかを明らかにするために、このプロセスに関わる酵素やタンパク質に特異的な抗体などの研究ツールが非常に役立ちます。
細胞代謝
細胞代謝は複雑に並んだ生化学反応によって成され、主なものに生体分子の合成 (同化)、維持、分解 (異化) があります。これらの総計が細胞のエネルギー状態を規定しています。このような代謝過程には様々な酵素やコファクターが関与し、細胞の基本的な構成要素となる脂質、アミノ酸、炭水化物、ヌクレオチドなどを合成、分解します。細胞代謝を成す様々な反応が、細胞の構成成分やエネルギーのプールに分子を加えたり差し引いたりするすべてのプロセスを包括しています。栄養素の取り込みが主な細胞代謝反応に影響します。そのため脂質、炭水化物、アミノ酸、ヌクレオチドなど、細胞内の主要な栄養素の代謝系を理解することが大変重要です。
脂質代謝
脂質 (脂肪) はエネルギーが豊富な栄養素で、細胞機能を支持するための燃料として利用されます。さらに、脂質は主要な細胞構造 (膜など) の重要な構成成分でもあり、多くの重要なシグナル伝達ネットワークにも関与します。細胞の種類によっては (脂肪細胞など) 脂質の貯蔵に特化しているものもあります。この脂質は栄養やエネルギーが欠乏した場合に生命機能を支持するため、異化作用を介して利用されます。中性脂肪やコレストロールなど主要な脂質は食事で摂取され、適切に消化吸収される必要があります。
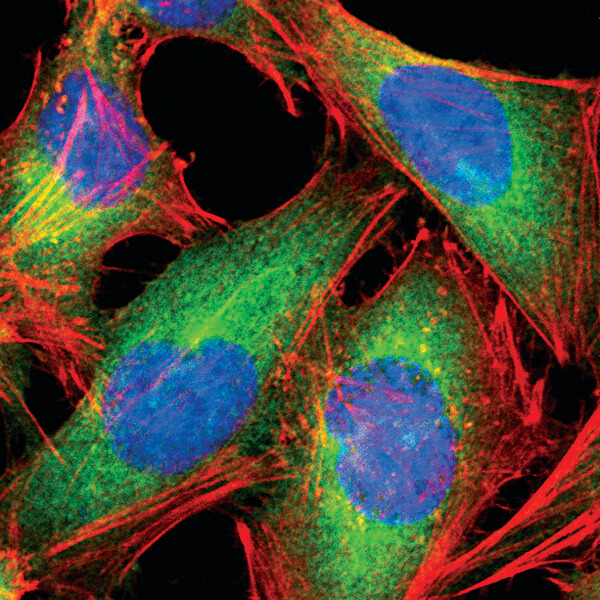
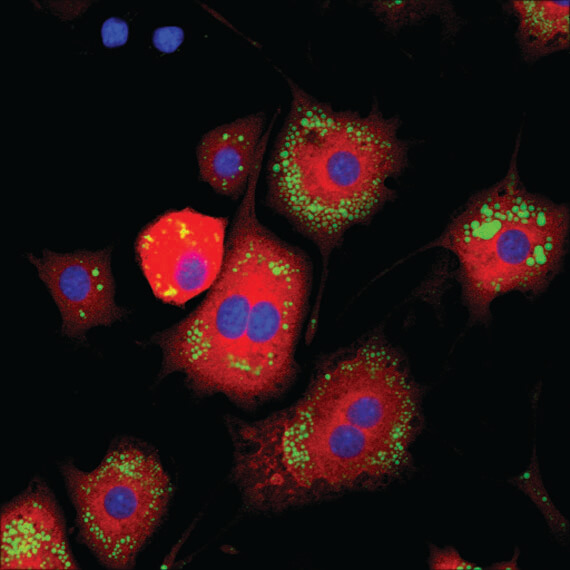
Fatty Acid Synthase (C20G5) Rabbit mAb #3180: HeLa細胞を#3180で染色 (緑) し、共焦点点IF解析しました。アクチンフィラメントをDY-554 phalloidin #13054 (赤) で染色しました。DRAQ5 #4084 (蛍光DNA色素) は、青の疑似カラーで示しています。
炭水化物代謝
炭水化物は、炭素、水素、酸素 (CHO) から成る「糖」であり、単純な単量体 (単糖、グルコースなど)、より複雑な二糖類 (スクロースなど)、高度に複雑な多糖類 (デンプンなど) といった形で存在します。炭水化物の代謝は摂食後速やかに開始され、酵素 (唾液アミラーゼなど) の放出によって多糖がより単純な糖へ分解されていきます。炭水化物の消化は小腸まで継続し、ここで膵臓アミラーゼが単糖への分解を完了します。グルコースはヒトの食事に含まれる主な単糖で、日々必要なエネルギーの大半がここから提供されています。
炭水化物代謝は、唾液アミラーゼとして知られる口腔内の酵素で始まります。唾液アミラーゼは複雑な炭水化物 (多糖) をより単純な分子に分解します。炭水化物の消化は小腸まで継続し、ここで膵臓アミラーゼが、部分的に消化された多糖をさらに最も単純な形 (単糖) まで分解します。グルコースは体が必要とする燃料の大半を供給する最も基本的な単糖です。グルコースは小腸で吸収されて血流に乗り、循環器系によってすべての器官に輸送されます。グルコースの細胞への取り込みはインスリンを介して誘導されます。グルコースは特に肝臓と筋細胞で盛んに取り込まれ、ここで代謝酵素がグルコースを多糖 (グリコーゲン) に変換します。この過程をグリコーゲン生成 (glycogenesis) と呼びます。グリコーゲン生成は、重要なエネルギーの貯蔵機能です。血中のグルコース濃度が低下すると、貯蔵されたグリコーゲンは迅速に加水分解されてグルコースになり、簡単に利用できるエネルギー源 (グルコース) を体に供給します。グリコーゲンは複数のグルコースのサブユニットから成ります。これは緊急時の燃料の供給源として機能し、血中グルコース濃度が低下すると速やかに分解されます。このグリコーゲンの分解を、グリコーゲン分解 (glycogenolysis) と呼びます。炭水化物の代謝経路の酵素機能の変化は、糖尿病や様々なグリコーゲン貯蔵疾患を引き起こします。
ヌクレオチドは核酸の構成単位で、糖 (五炭糖) とリン酸、塩基から成ります。 RNA、DNAの合成には構造のよく似たヌクレオチドが使用されますが、塩基部分に大きな違いがあり、RNA合成ではチミジンの代わりにウラシルが使われます。また、五炭糖にはリボース、デオキシリボースの2種類があり、それぞれRNA、DNAの合成に使われます (省略されることが多いですが、RNAはリボヌクレオチド、DNAはデオキシリボヌクレオチドによって構成されます)。デオキシリボースの合成にはリボースからの-OH基の除去 (還元反応) が必要であり、この反応を触媒するリボヌクレオチドレダクターゼがDNA合成の律速となります。重要なことに、DNAとRNAの分解は細胞で継続的に起こっています。DNAとRNA分解で生じるプリンとピリミジン産物は、将来の核酸合成のためにリサイクルされるか、廃棄物として除去されます。様々なタイプの核酸の周期的な分解と合成は、エネルギーの貯蔵と生成に重要であり、細胞の恒常性維持に重要です。
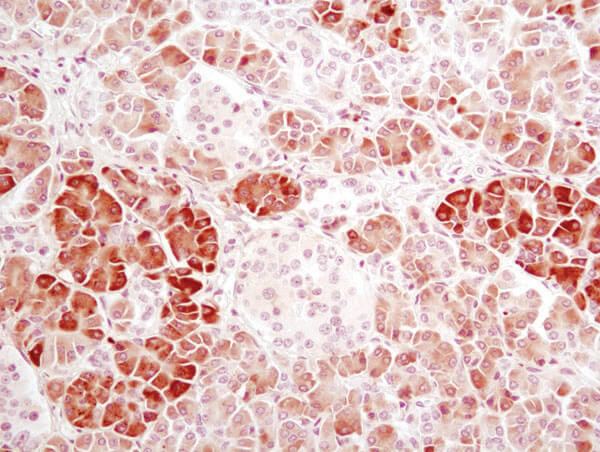
α-Amylase (D55H10) XP® Rabbit mAb #3796:α-Amylase (D55H10) XP® Rabbit mAb #3796を用いたパラフィン包埋ヒト膵臓のIHC解析。
アミノ酸代謝
タンパク質は非常に多くの機能を担う分子で、その一部に1) 細胞内外の構造 (細胞/組織/器官) の構成成分として機能する、2) シグナル伝達を媒介する、3) 分子輸送 (酵素としてこれらの制御に関与する)、4) 免疫機能を媒介する等々があります。アミノ酸はタンパク質の構成単位で、これが連結してポリペプチド鎖を形成します。これが折り畳まれて三次元構造が完成すると成熟タンパク質といて機能します。タンパク質は細胞の構成物質の大きな割合を占め、燃料などとしても利用される重要な栄養素でもあります。
タンパク質の消化は胃で始まり、胃酸と消化酵素ペプシンがタンパク質をより単純なポリペプチドに分解します。このようなポリペプチドはその後さらにアミノ酸まで分解されます。これは他のタンパク質の材料としてリサイクルされるか、肝臓でさらにα-ケト酸に分解されます。α-ケト酸はエネルギーの産生や、グルコース、脂肪、新しいアミノ酸の合成に利用することができます。アミノ酸の分解では、アンモニウムイオンが産生されます。アンモニア毒性を除去する尿素サイクルの機能については後記します。また、タンパク質は飢餓条件で非常に有用なエネルギー源となることも重要です。タンパク質の分解により、クエン酸サイクルに送り込まれる代謝中間体が産生されます (後記)。
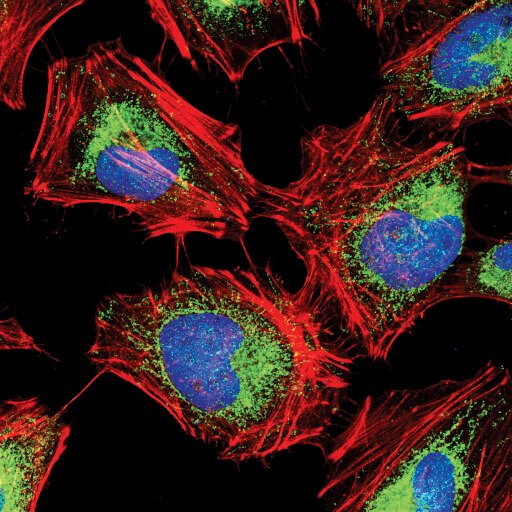
Hexokinase I (C35C4) Rabbit mAb #2024:Hexokinase I (C35C4) Rabbit mAb #2024 (緑) を用いたHeLa細胞の共焦点IF解析。アクチンフィラメントはDY-554 phalloidin #13054 (赤) で染色しました。DRAQ5 #4084 (蛍光DNA色素) は、青の疑似カラーで示しています。
ヌクレオチド代謝
ヌクレオチドは、プリンとピリミジンの2つの主なグループに分類されます。どちらもリン酸基と五炭糖から成り立っていますが、大きさと窒素塩基が異なります。
アデニンとグアニンはプリンであり、シチジン、ウリジン、チミジンはピリミジンです。プリンとピリミジンの合成のエネルギー源としてアデノシン三リン酸 (ATP) やグアノシン三リン酸 (GTP) が利用されます。ATPとGTPは細胞の主なエネルギー担体で、リン酸基どうしの結合にエネルギーが蓄えられています。リン酸基同士の結合が一つ切断される際に放出されるエネルギーが、様々な細胞機能を駆動するためのエネルギーとして利用されます。リン酸基はホスファターゼ酵素を介して端から切除され、ATPとGTPからアデノシン二リン酸とグアノシン二リン酸 (ADP、GDP)、さらにアデノシン一リン酸とグアノシン一リン酸 (AMP、GMP) が産生されます。これは可逆的な反応で、リン酸基はキナーゼとして知られる酵素によりADP/GDPやAMP/GMPに付加されます。ピリミジン合成経路ではウリジンからUTP、シチジンからCTPヌクレオチドも合成されます。
ミトコンドリアと代謝におけるミトコンドリアの機能

ミトコンドリア:ミトコンドリアのランドスケープ
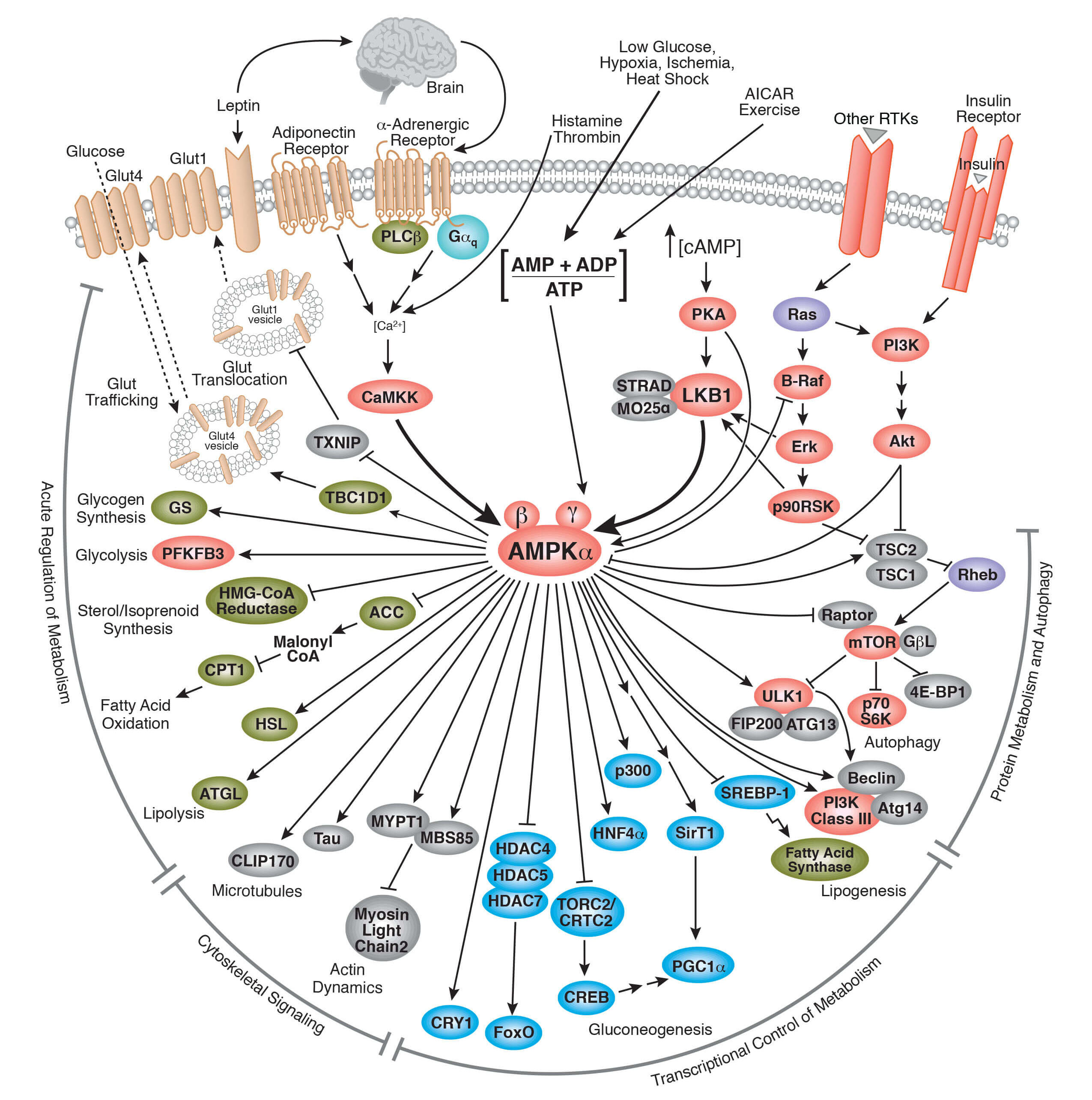
ミトコンドリアは、細胞代謝とエネルギー産生で中心的な役割を果たす細胞内小器官です。この動的な細胞小器官は、異なる代謝的な入力を統合し、主要な代謝経路と細胞運命決定機構をコントロールしながら、投入された栄養素をエネルギー通貨、主にATPに変換する効率的なエンジンとして機能します。栄養素の代謝産物からのATPを合成することが、ミトコンドリアの機能の重要な側面と考えられています。ミトコンドリア内膜の呼吸鎖サブユニットを通して中間産物の電子の受け渡しをすることで、クエン酸回路の代謝産物からATPが合成されます。電子の受け渡しによってプロトンが膜間腔に汲み出され、ミトコンドリア内膜の内外に生化学的なプロトン濃度勾配が形成されます。プロトンの汲み出しは、ミトコンドリア内膜にある主要な呼吸鎖複合体I、III、IVによって行われます。これらは電子の受け渡しで得られるエネルギーを用いてプロトンをミトコンドリアマトリクスから膜間腔に汲み出すプロトンポンプとして機能します。酸素分子最終的な電子の受容体で、複合体IVから電子を受け取りH2Oに還元されます。こうして (ミトコンドリア内膜を挟んで) ミトコンドリアマトリクスと膜間腔の間に生み出された電気科学的勾配 (プロトン濃度勾配) が、ADPからATPを産生するエネルギーを供給します。この効率的なエネルギー産生プロセスは、酸素に依存することから「好気的呼吸」と呼ばれます。好気的呼吸は酸素を必要としない嫌気的代謝 (グルコースから乳酸を産生する嫌気的解糖など) などと比較して、最初の栄養素の投入に対し最も効率的にエネルギー収益 (ATP) を生み出します。ミトコンドリアの機能とエネルギーの生成は、脳内の細胞 (ニューロン)、心臓 (心筋細胞)、膵臓 (β島) など、体内のエネルギーの要求が高い組織と細胞で特に重要です。ミトコンドリアの機能は全細胞にとって重要で、ミトコンドリアの機能に障害があると疾患の原因となります。ミトコンドリア機能障害の結果として生じる疾患の重篤度は、それが生じた細胞の代謝要求性によって感受性が異なります。ミトコンドリア機能障害に起因する様々な代謝疾患は、セクション4で総説します。
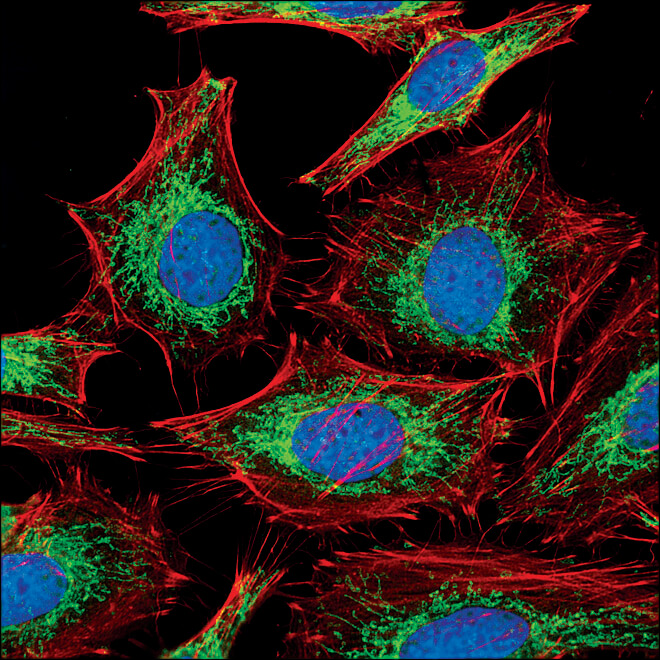
COX IV (3E11) Rabbit mAb #4850:COX IV (3E11) Rabbit mAb #4850 (緑) で染色したHeLa細胞の共焦点IF解析。アクチンフィラメントをAlexa Fluor® 555 phalloidin #8953 (赤) で染色しました。DRAQ5 #4084 (蛍光DNA色素) は、青の疑似カラーで示しています。
ミトコンドリアは独自の生理機能を持つことでも知られ、細胞の代謝に寄与しています。しかしながら、ミトコンドリアは細胞内に単独で存在しているわけではなく、細胞質全体の動的なネットワークに組み込まれています。このネットワーク内のミトコンドリアのライフサイクルには、生合成、融合と分裂、運動、分解といったプロセスがみられます。生合成は新しいミトコンドリアの発生です。融合は別々のミトコンドリアが合併して1つになることです。分裂はその逆で、1つのミトコンドリアが別々のミトコンドリアに分離することです。これらのプロセスは、ミトコンドリアが細胞の様々なエネルギー要求に対応するため、また、ミトコンドリアの損傷に反応して細胞のミトコンドリアの機能を保存するために、継続的に発生しています。ミトコンドリアの運動は、ミトコンドリアがその機能が必要とされている場所に移動するプロセスです。最後に、マイトファジーはミトコンドリアに選択的に影響を及ぼすオートファジーに似たプロセスで、ミトコンドリアの構成成分が分解されます。オートファジーは特に、損傷したミトコンドリアの除去を行います。ミトコンドリアのオートファジーやマイトファジー機構の障害は、機能不全のミトコンドリアの蓄積の原因となります。パーキンソン病の発症など、いくつかの疾患でマイトファジーの障害が関与することが分かっています。これらのことから、ミトコンドリアの機能の制御と維持は、代謝恒常性の維持に非常に重要であると言えます。
主要な代謝経路
以下に、細胞が利用している主要な代謝経路について解説します。いずれの経路も、特定の化学反応を触媒する酵素が中心的な役割を担っています。このような酵素を介して代謝基質を別分子に変換しますが、その過程で複数の中間ステップを経て様々な代謝産物を産生します。
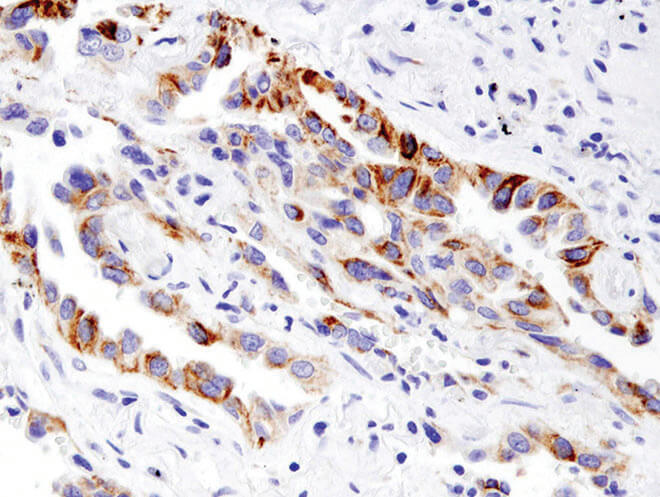
BNIP3 (D7U1T) Rabbit mAb #44060:BNIP3 (D7U1T) Rabbit mAb #44060を用いた、パラフィン包埋ヒト肺がんのIHC解析。
同化、異化、両性代謝
マクロファージは損傷した細胞や外来物質を認識して活性化され、これらを排除します。マクロファージはほとんどの組織に存在し、必要に応じて感染や死細胞に対処します。これがこの細胞の名前の由来です (ギリシャ語で「大食らい」の意味)。マクロファージは局在する部位によって様々な形をとり、貪食以外の追加機能を果たします。
組織の損傷や感染が起こると、血中の単球が患部に動員されて分化し、マクロファージとなります。組織局在性によって異なるタイプのマクロファージがあります。例えば肝臓のクッパー細胞、肺の肺胞マクロファージ、脳のミクログリアなどです。これらの異なるタイプのマクロファージは全て単球由来ですが、常在する組織によって機能が特殊化します。一般的な貪食のほとんどは、組織常在型のマクロファージに実行されます。死細胞や異物の貪食のほか、マクロファージはサイトカインを介して他の免疫細胞にシグナルを伝えます。マクロファージはある程度、抗原提示の機能を持ち、これによってT細胞と協調して適応免疫をサポートします。また、マクロファージはIL-12などのサイトカインを分泌して局所免疫反応に重要な役割を果たします。一方で他のマクロファージはIL-10を大量に産生し、組織修復に関与しています。このように、「大食らい」は貪食という主な仕事に加え、免疫系の様々な機能を担っています。
解糖系
解糖系は、複数の生化学的反応を経てグルコースをピルビン酸とATPへ分解する経路です。解糖系の反応は細胞質で起こり、一連の酵素を介して6個の炭素を持つグルコース分子を、3個の炭素を持つピルビン酸分子2個に分解します。解糖系は酸素を必要としないため、嫌気性生物で支配的な異化反応です。解糖系は酸素の供給が制限された環境では、好気性生物にも有用な経路です。
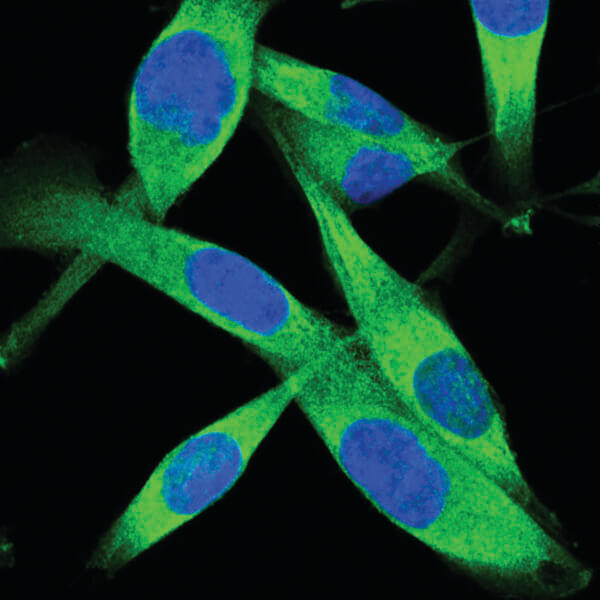
PKM2 (D78A4) XP® Rabbit mAb #4053:PKM2 (D78A4) XP® Rabbit mAb #4053 (緑) を用いたA204細胞の共焦点IF解析。DRAQ5 #4084 (蛍光DNA色素) は、青の疑似カラーで示しています。
クエン酸回路 (クレブス回路)

クレブス回路:クレブス回路のタンパク質
クエン酸回路はクレブス回路としても知られる経路で、解糖系に続く細胞内グルコース代謝の次のステップです。この反応はミトコンドリアで起こり、解糖系の最終産物ピルビン酸の誘導体であるアセチル補酵素A (アセチルCoA) により開始されます。クエン酸回路は酸素を必要とし、二酸化炭素と水を副産物として産生します。この経路は高エネルギー分子のNADH、FADH2、ATPを産生する、一連の酸化還元反応で構成されています。解糖系ではグルコース1分子からそれぞれに2分子のピルビン酸が産生されます。したがって、クエン酸回路2周駆動され、1周当たり二酸化炭素が2分子、NADHが3分子、FADH2が1分子、ATPが1分子ずつ産生されます。クエン酸回路そのものはそれほどの量のATPは産生しませんが、細胞の重要なエネルギー通貨であるNADHとFADH2が産生されます。これらは酸化的リン酸化に共役した電子伝達系の電子担体としての役割を果たし、これによって高エネルギーが産生されます。
酸化的リン酸化
細胞呼吸 (糖代謝) の最後の段階は、酸化的リン酸化 (OXPHOS) です。OXPHOSはミトコンドリアの内膜を挟んで起こります。一連の酸化還元反応において、ミトコンドリア内膜にある5つの膜貫通型酵素複合体が電子を1つの分子から別の分子に移動させます。これは電子伝達系として知られています。電子の受け渡しで得られるエネルギーを利用してプロトンポンプを駆動し、ミトコンドリア内膜を横切ってミトコンドリアマトリクスから膜間腔へプロトンを輸送し、プロトンの濃度勾配を形成させます。これが、OXPHOSを生じさせる電気化学的勾配の起源です。膜間腔に蓄積したプロトンはその後、電子伝達系の最後の複合体であるATP合成酵素を通って、形成された濃度勾配に従ってミトコンドリアマトリクスに流れ込みます。ATP合成酵素分子は「分子モーター」として機能し、プロトンが濃度勾配を流れ落ちることで得られるエネルギーを利用して、ADPにリン酸基を付加する反応を触媒します (これによってATPが産生されます)。酸化的リン酸化によって、グルコース1分子当たり30 - 36分子のATPが産生されます。
ペントースリン酸経路
上記とおり、細胞呼吸では解糖系で産生されたピルビン酸がクエン酸回路と酸化的リン酸化に供されます。しかし、解糖から分岐し、DNAとRNAの合成に必要な糖を産生する、別の細胞質経路があります。ペントースリン酸経路は、解糖系の最初のステップで産生される分子、グルコース-6-リン酸を利用します。グルコース-6-リン酸は、グルコースにリン酸基が付加したものです。ペントースリン酸経路は、NADPH、5個の炭素を持つ糖 (五炭糖) ペントース、ヌクレオチド合成の前駆体分子リボース-5-リン酸を産生します。NADPHは、脂肪酸代謝や活性酸素種 (ROS) の制御など、他の生合成過程でも重要な機能を果たします。
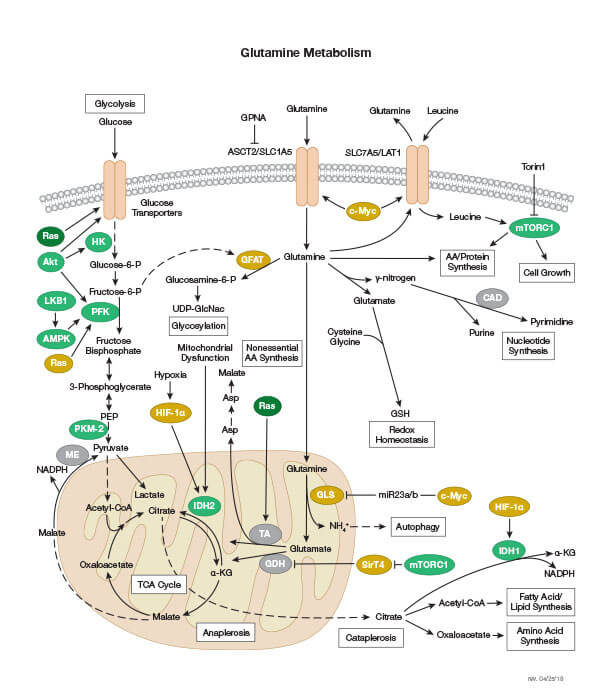
グルタミン代謝
グルタミンは、急速に増殖している細胞にとって重要な燃料源です。特異的なアミノ酸輸送体を介して細胞内に輸送され、ミトコンドリアでグルタミン酸に変換されます。グルタミン酸はさらに、クエン酸回路の中間産物であるα-ケトグルタル酸に変換されます。
尿素回路
尿素回路はオルニチン回路とも呼ばれ、体内のアンモニアの毒性の蓄積を防ぐために必要な経路であり、主に肝臓で起こります。この回路は、アミノ酸分解の副産物であるアンモニウムイオンから尿素を産生する生化学反応で構成されています。この回路では、タンパク質代謝中に起こるアミノ酸のアミノ基転移で産生されたアンモニアに二酸化炭素を加え、最終的に尿素と水を産生します。これらはその後、腎臓で尿として排泄されます。尿素回路の最初のステップはミトコンドリアで起こり、後のステップは細胞質に移って行われます。
脂肪酸合成
脂肪酸は、細胞内のエネルギー源でもあり貯蔵先でもあります。さらに、脂肪酸は細胞のシグナル伝達においても重要な役割を果たし、細胞機能に大きな影響を与えます。脂肪酸合成は細胞質で起こり、アセチルCoAとNADPHから脂肪酸を合成します。このプロセスは脂肪酸シンターゼによって触媒されます。脂肪酸合成に必要なアセチルCoAは、解糖系によるグルコースの分解で供給されます。グルコース分解は、グリセロールも産生します。これは3個の脂肪酸サブユニットと結合して中性脂肪 (トリグリセリド) ができます。リン脂質の生成も脂肪酸代謝の重要な側面であり、リン脂質は生体膜の主要な構成成分となります。グリセロールにリン酸基と脂肪酸2個が結合するとリン脂質が形成されます。リン脂質は細胞内で多数の機能を持ちますが、最も重要なものは細胞膜を成す脂質二重層の形成です。
リン脂質は細胞膜やオルガネラ膜の構成成分ですが、医薬品の合成にも利用されています。これは、生体膜の透過性を向上させ、生物学的利用効率を改善するためです。
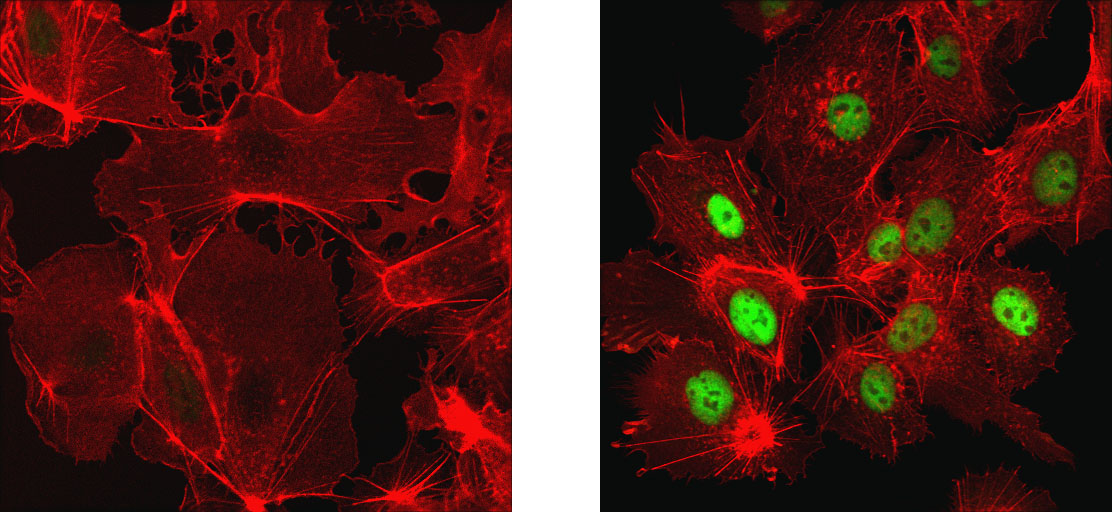
Lipin 1 (D2W9G) Rabbit mAb #14906:分化誘導 (7日間) した3T3-L1細胞をLipin 1 (D2W9G) Rabbit mAb #14906 (赤) を用いて染色し、共焦点IF解析を行いました。脂肪滴をBODIPY® 493/503 (緑) で標識しました。DRAQ5 #4084 (蛍光DNA色素) は、青の疑似カラーで示しています。
脂肪酸のβ酸化
脂肪酸のβ酸化は、脂肪酸を分解してその構成成分であるアセチルCoAを取り出すプロセスであり、ミトコンドリアで起こります。アセチルCoAはこの後クエン酸回路に入り、酸化反応とNADH、FADH2の産生が行われます。
糖新生
糖新生は炭水化物以外の物質からグルコースを合成する反応です。グリコーゲン分解と同様、血糖値の低下を抑える適応プロセスであり、主に肝臓で起こります。糖新生は通常、栄養素の摂取が低い時、激しい運動を行った時、あるいは低炭水化物食品を摂取している時に起こります。
一炭素代謝
一炭素代謝は、アミノ酸やヌクレオチドなどいくつかの分子の同化に必要な、葉酸依存性代謝経路の1つのグループです。この経路では葉酸が一炭素基の担体の役割を果たし、ドナー分子から一炭素基の除去と移送を促進しています。一炭素基の輸送に利用される分子には次の3つがあります。1) 葉酸誘導体でいくつかの酵素のコファクターとして機能する、テトラヒドロ葉酸、2) メチル基の供与体である、s-アデノシルメチオニン (SAM)、3) メチル化と炭素再構成反応の補酵素であるビタミンB12。アミノ酸とヌクレオチド合成における役割の他に、この経路の代謝物であるSAMは共通のメチル基供与体として機能するため、DNAやヒストンのメチル化制御にも重要です。
酸化ストレスとその代謝における役割
ミトコンドリアの電子伝達系を介したATP産生の副産物として、活性酸素種 (ROS) が産生されます。これは反応性の高い分子で細胞小器官やその他の細胞構造に酸化的損傷を起こします。細胞は抗酸化システムを備えており、ROSの低い生理的レベルではその毒性をスーパーオキシドジスムターゼ (SOD)、グルタチオン、カタラーゼなどによって十分に制御できます。現在では、基底レベルのROSが細胞の生理的なパスウェイに重要な機能を持つことがわかっています。しかしながら、病的なレベルのROSがタンパク質、脂質、DNAに損傷を与えることも明らかにされており、これがミトコンドリア代謝を妨げたり、細胞機能や生存に悪影響を及ぼしたりすることがあります。様々な細胞ストレス環境下で細胞機能や生存を維持するため、ROSによるミトコンドリア損傷に効率的に対処することが重要です。
酸化ストレスとは
様々な環境条件や細胞機能の撹乱が酸化ストレスの原因となります。代謝に注目すると、エネルギーの需要に対して栄養素が過剰に供給された場合に酸化ストレスが生じます。栄養素の過剰供給によって電子伝達系が促進されますが、これは同時に呼吸鎖複合体からの電子の「漏れ」の原因となります。こうして漏れ出した電子がO2と反応してROSを産生します。呼吸鎖構成成分の機能障害も電子伝達の撹乱の原因となり、ROSレベルを増加させます。同様に、抗酸化酵素の発現低下や機能障害はROSの蓄積の原因となります。ミトコンドリアがROSの主な発生源ですが、この他、キサンチンオキシダーゼや食細胞にみられるシトクロムP450オキシダーゼ系などで産生されるROSや活性窒素種 (PNS) も酸化ストレスの原因となります。
酸素種 (ROS)
ROSには様々な形があり、フリーラジカル (不対電子を持つもの) と非ラジカル (不対電子を持たないもの) に大別されます。自由電子が最初にO2と反応すると、スーパーオキシドアニオン (O2-) を形成します。これは反応性が非常に高く、不安定なROSとして知られています。スーパーオキシドは速やかにスーパーオキシドジスムターゼ (SOD) によって過酸化水素 (H2O2) に変換されます。過酸化水素は比較的安定していますが、遷移金属と相互作用することでフェントン反応として知られる過程を経て有害なヒドロキシラジカル (・OH) に変換されます。ヒドロキシラジカルは最も反応性の高いROSで、細胞内タンパク質と脂質に最も酸化的損傷を引き起こしやすいものです。
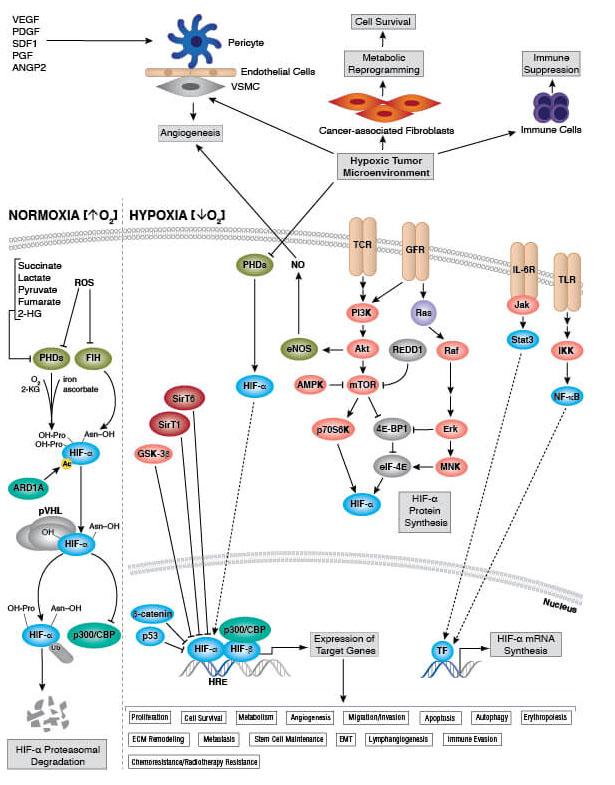
低酸素と細胞呼吸
低酸素 (酸素に制限がある) 条件下では電子伝達系は正常に進行しますが、最終的な電子受容体となる酸素が不足します。これを放置すると電子が漏れ出し、ROSの産生が増加する原因となります。このため、細胞は酸化ストレスに関連して細胞呼吸に影響を及ぼす特殊な低酸素応答パスウェイを進化させてきました。これらのパスウェイは代謝活性を抑制して生体エネルギー機構が空回りするのを防いでいます。低酸素応答の制御で中心的な役割を担うのが、転写因子HIF-1 (hypoxia-inducible factor-1) です。HIF-1は酸素量の低下を感知して電子伝達系、タンパク質の翻訳 (大量のATPを必要とするプロセス)、Na-K-ATPaseの活性を抑制します。このような統制された反応によって、細胞とミトコンドリアは低酸素条件に耐え、正常な酸素レベルに戻るまで持ちこたえます。
HIF-1α (D1S7W) XP® Rabbit mAb #36169:塩化コバルト処理 (500 μM、24時間) したHep G2細胞 (右) と未処理コントロール細胞 (左) を HIF-1α (D1S7W) XP® Rabbit mAb #36169 (緑) を用いて染色し、共焦点IF解析しました。アクチンフィラメントをDyLight™ 554 Phalloidin #13054 (赤) で染色しました。
酸化ストレスのバイオマーカー
酸化ストレスは多面的で複雑なプロセスであるため、細胞や組織内の絶対的なバイオマーカーを定義するのは容易ではありません。
1つの方法として、酸化的損傷の結果として生じる標的分子 (タンパク質など) の生化学的修飾を調べる方法があります。例えば、タンパク質カルボニル化アッセイでは、DNPH (2,4-dinitrophenylhydrazine) を用いて活性酸素に修飾されたタンパク質を検出します。特に循環器疾患では、酸化された低密度リポタンパク質も一般的なバイオマーカーとして利用されています。酸化的損傷によって脂質の修飾もみられます。よく利用されているのは、脂質の過酸化に応答して産生される4-HNE (4-hydroxynonenal) です。また、細胞の抗酸化システムの活性も利用することができ、例えば、内因性グルタチオンレベルは酸化ストレスの指標となります。DNAとRNAもROSの標的となるので、これらヌクレオチドの酸化も酸化ストレスのバイオマーカーとして使用できます。Nrf2やHIF-1など、酸化ストレスに応答する既知のパスウェイに着目して酸化ストレスを推定することもできます。酸化ストレスのバイオマーカーの探求は常に進化しているので、直近の文献を確認してから目的に合致したアッセイを決定することが求められます。
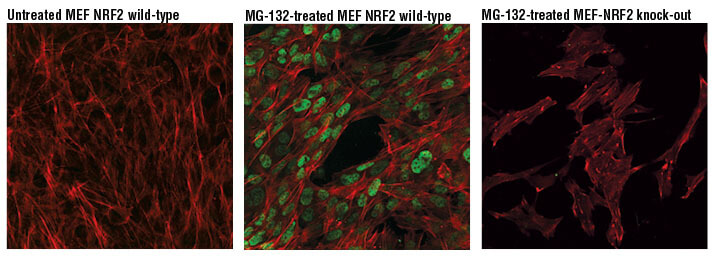
NRF2 (D9J1B) Rat mAb (IF Specific) #14596: MG-132 #2194処理 (10 μM、8時間) した野生型MEF (中央) と未処理コントロールMEF (左)、同様にMG-132 #2194処理 (10 μM、8時間) したNRF2ノックアウトMEFを調製しました。これらをNRF2 (D9J1B) Rat mAb (IF Specific) #14596 (緑の疑似カラー) を用いて染色し、共焦点IF解析しました。アクチンフィラメントをAlexa Fluor® 488 Phalloidin #8878 (赤の疑似カラー) で染色しました。
代謝疾患
細胞代謝の恒常性を維持することは非常に重要であり、代謝経路の障害が様々な代謝疾患を引き起こします。以下に、主な代謝疾患とその原因をまとめました。
代謝疾患の原因
代謝疾患は遺伝子変異によって発症する場合と、環境要因によって消化器官や代謝機能をもつ器官 (肝臓や膵臓など) の機能が障害を受けて発症する場合があります。
代謝疾患の種類
米国国立衛生研究所は現在、代謝疾患を次のように分類しています。
- 酸と塩基の不均衡
- 代謝性脳疾患
- カルシウム代謝の疾患
- DNA修復欠損性疾患
- グルコース代謝疾患
- 高乳酸塩血症
- 鉄代謝疾患
- 脂質代謝疾患
- 吸収不良症候群
- 代謝症候群X
- 先天性代謝異常症
- ミトコンドリア性疾患
リン酸代謝疾患
- ポルフィリン症
- タンパク質恒常性の喪失
- 代謝性皮膚疾患
- 消耗症候群
- 水電解質不均衡
主な代謝疾患
代謝疾患の大部分を占め、人々の生活に影響を与えている疾患があります。糖尿病/インスリン抵抗性、肥満/代謝症候群 (メタボリックシンドローム)、循環器疾患、腎組織障害、先天性代謝異常などがこれに当たります。
がんと代謝
がん細胞生物学における代謝の重要性が増し続けています。初めにがんと代謝の関係性が理解しされ始めたのは、がん細胞がエネルギー効率の良い好気的呼吸ではなく、効率の悪い発酵に強く依存するというOtto Warburg博士の研究成果です。この「ワールブルグ効果」は、多くのがん細胞にみられる特性であり、増殖と生存を促進するための代謝適応を反映していると考えられます。この代謝適応の意義については現在、低酸素条件下で細胞の構成成分 (タンパク質、脂質、核酸など) を安定して産生することで、早く無秩序な細胞増殖が促進されるため、正常細胞に比べてがん細胞の生存が有利になる、という仮説が有力です。別の研究では、盛んに増殖するがん細胞は、栄養素を細胞の構成成分の産生に充てる必要があるため、異化 (栄養素の消費) から同化 (生体高分子の構築) に代謝経路を切り替えていることが示唆されています。ワールブルグ効果の他にもがん細胞の代謝にはいくつかの特徴がみられます。グルコースとアミノ酸摂取の障害、異なる方法による栄養素の獲得、解糖系/TCA回路の中間産物を利用した生合成とNADPHの産生、窒素要求性の亢進、代謝産物による遺伝子制御の変化、微小環境との代謝的相互作用などがこれに当たります。
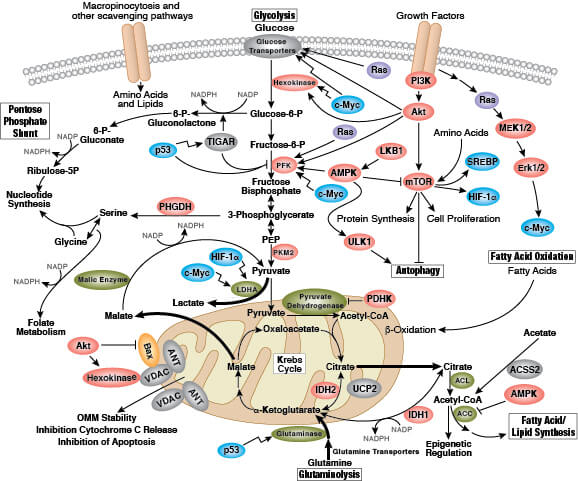
糖尿病/インスリン抵抗性
糖尿病は血糖値が生理的に健常なレベルを慢性的に超えることで発症します。糖尿病には1型糖尿病と2型糖尿病の二種類があります。1型糖尿病は通常幼児期から成人早期 (思春期を含む) に発症し、血中グルコースの細胞への取り込みを誘導するホルモンであるインスリンが十分に産生されなくなります。自己免疫などによる膵島のβ細胞 (インスリン産生細胞) の障害がこのタイプの糖尿病の原因となります。2型糖尿病では、インスリンが作られない場合と、体がインスリンの存在に対し脱感作 (すなわちインスリン抵抗性) されている場合があります。2型糖尿病は年齢に関係なく発症し、多くの場合食生活に起因します、1型糖尿病と2型糖尿病の患者は、心臓病、腎臓病、歯科疾患、循環器異常、眼疾患および神経損傷を併発することが多くあります。
肥満
肥満は、皮下や内臓脂肪として貯蔵される過剰な脂肪組織に起因します。肥満は通常、体格指数 (BMI:Body-mass index) をもとに診断されます。体重を身長の関数として算出し、性別や年齢にしたがって判定します。肥満は米国で最も高頻度に見られる疾患の1つで、その驚くべき統計値は年々増加の一途を辿っています。肥満は、心臓病や循環器疾患、2型糖尿病、がんなど、様々な疾患を高頻度に併発する、予防可能な早逝の原因の1つです。遺伝的に肥満になりやすい人もいますが、肥満の症例の大半は食生活や生活習慣に起因します。
代謝症候群
代謝症候群は肥満に関連するもう1つの疾患です。これは代謝疾患を併発した合併症であり、さらに糖尿病、心臓病、脳卒中などを高頻度に併発します。代謝症候群の診断は、患者が次の症状のうち3つに該当する場合に下されます:腹部肥満、高トリグリセリド (中性脂肪) 血症、低HDLコレステロール (善玉コレステロール) 血症、高血圧、空腹時の高血糖。
心血管疾患
アメリカ心臓協会は、循環器疾患を次の1つ以上の症状として定義しています:心臓病、心臓発作、脳卒中、心不全、心臓弁の問題。肥満と糖尿病は循環器疾患のリスクを上昇させます。さらに、エネルギーへの依存度が高いため、心細胞が細胞内代謝経路に障害を起こしたとき、代謝症候群/肥満とは独立して循環器疾患を発症することがあります。
腎組織障害
代謝症候群は、慢性腎臓病とも関連しています。様々な臨床的特徴が代謝症候群との関連性を示し、代謝症候群腎機能障害や組織損傷に寄与する考えられています。これには高インスリン血症、レニン-アンジオテンシン-アルドステロン系の活性化、酸化ストレスの上昇、炎症性サイトカイン産生などがあります。
先天性代謝異常
先天性代謝異常は、代謝パスウェイに関与する主要な酵素の機能不全または欠損によって起こる、遺伝性の希少疾患です。以下にライソゾーム病、アミノ酸代謝疾患、炭水化物代謝疾患について解説します。
ライソゾーム病
ライソゾーム病は、ライソゾームの主要な酵素の機能障害によって、本来ライソゾームの標的として分解されるべき物質が蓄積する疾患の総称です。ライソゾーム病の例として、ゴーシェ病とニーマン・ピック病が挙げられます。ゴーシェ病は、グルコセレブロシダーゼの機能障害に起因し、スフィンゴ脂質であるグルコセレブロシドが白血球やマクロファージに蓄積します。ニーマン・ピック病は脂質貯蔵の異常がみられる疾患で、脾臓、肝臓、肺、骨髄、脳に高レベルの脂質が蓄積します。ニーマン・ピック病は、遺伝的基盤と関連症状に基づいて4種類に分類されます。
アミノ酸代謝異常症
アミノ酸代謝異常症は、アミノ酸の異化作用に異常がみられる遺伝疾患です。アミノ酸代謝異常症の例として、フェニルケトン尿症とメープルシロップ尿症が挙げられます。フェニルケトン尿症は乳児に発症し、アミノ酸であるフェニルアラニンとチロシンの分解に障害が起こります。このため血中にフェニルアラニンの蓄積が起こります。フェニルアラニンは脳に障害を引き起こし、フェニルケトン尿症が治療されず放置されると知的障害の原因となる発育不全を引き起こします。同様に、メープルシロップ尿症も幼児に発症し、ロイシン、イソロイシン、バリンの分解に障害が起こります。この結果、アミノ酸副産物が蓄積して体液がメープルシロップのようなにおいを放つようになり、最終的には神経機能障害を引き起こします。
糖代謝異常症
糖代謝異常症は、糖代謝に関与する酵素の機能不全に起因します。糖原病はグリコーゲンの生化学的プロセシングに関与する主要な酵素の変異に起因する遺伝疾患です。糖原病には10種以上のタイプがあり、障害の起こった酵素に基づいて分類されます。障害の起こった酵素の組織分布によって様々な器官が影響を受けるため、様々な症状がみられます。炭水化物代謝疾患の別の例として、炭水化物の異化作用に関与するG6PD (glycogen-6-phosphate dehydrogenase) の欠損に起因するものが挙げられます。G6PDが欠損すると、活性酸素種の蓄積によって赤血球が溶解し、溶血性貧血の症状を引き起こします。