

BROMOタンパク質ドメイン
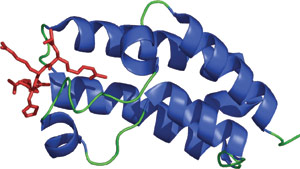
アセチル化リジン (赤) に結合しているGcn5pのブロモドメイン
ドメイン結合および機能
約110アミノ酸残基からなるブロモドメインは、酵母からヒトまでの100種類以上のタンパク質において同定され、アセチル化されたリジン残基に結合することが知られています。N末端およびC末端のヒストンテールのリジン残基のアセチル化は、リジンのメチル化や、セリン/スレオニンのリン酸化と並んで重要な翻訳後修飾で、クロマチン構造変化をさせ、遺伝子発現のエピジェネティックな制御に関与します。ブロモドメインは、ヒストンアセチルトランスフェラーゼや、ある種のヌクレオソームリモデリング複合体のATPaseサブユニットなど、クロマチンの構造や遺伝子発現を調節するタンパク質によくみられます。ブロモドメインはペプチド中に含まれるアセチル化されたリジンと相互作用することが知られている唯一のドメインですが、ブロモドメインによるアセチル化リジンの認識様式は、ヒストンアセチルトランスフェラーゼによるアセチルCoAの認識と同様です。ブロモドメインタンパク質によるアセチル化リジンの認識はヒストンに限りません。例えば、CBP (CREB binding protein transcriptional coactivator) のブロモドメインは、p53のアセチル化されたLys382を認識します。このブロモドメインとアセチル化p53の相互作用は、DNA損傷に続いて起こり、p53が誘導するCDK阻害剤のp21の転写活性化および細胞周期停止を促進します。
構造の参考文献
- Owen, D.J. et al. (2000) EMBO J. 19(22), 6141–6149.
ドメインタンパク質の例
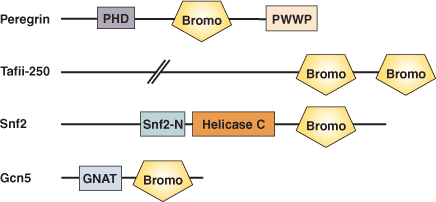
結合の例
BROMOドメインタンパク質 | 結合パートナー | 特異的な結合サイト |
PCAF | Tat | BSYGRKAcKRRQRC |
CBP (CREB Binding Protein) | 三元複合体因子Elk-1 | 未知、SSPQPKKAcKPLDGE |
| p53 | SHLKSKKAcGQSTSRHKK, SSPQPKKAcKPLDGE |
Gcn5p | Histone H4 | AKAcRHR |
Celtix-1 | IRF-2 | 高アセチル化型IRF-2 |