

細胞骨格の制御と小胞輸送
CSTのシグナル伝達パスウェイ図で個々のタンパク質名をクリックすると、それに関連した研究リソースや製品情報を検索することができます。さらに、教育・研究のために、パスウェイ図をダウンロードすることも可能です。
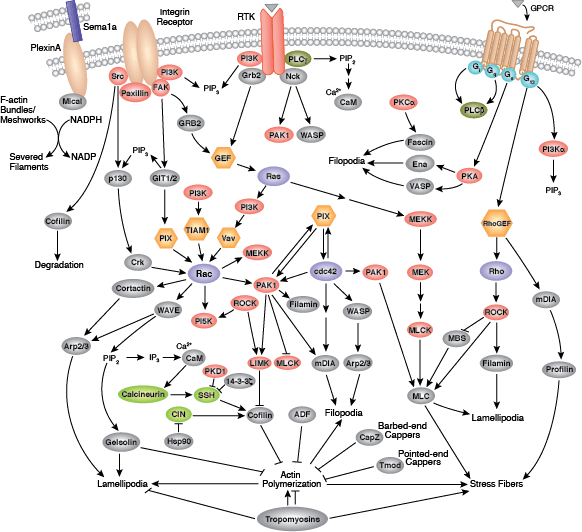
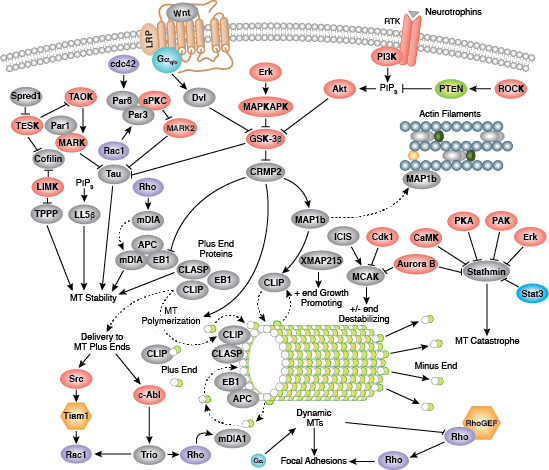

細胞は3つのタイプの細胞骨格繊維から構成されます。それは、微小管、マイクロフィラメント (アクチンフィラメント)、そして中間径フィラメントです。細胞骨格のシグナル伝達は、細胞分裂、接着、極性、遊走、繊毛や鞭毛を利用した移動など、細胞内の様々な重要プロセスを制御しています。
微小管は、球状のTubulinサブユニットから構成され、これはα/β-Tubulinヘテロ二量体から成り、すべての真核細胞に共通しています。γ−Tubulinは、Tubulinサブユニットの重合の核を形成して微小管重合体を形成するために必要です。多くの細胞の運動は、繊毛や鞭毛の運動、膜小胞の細胞質内輸送、神経細胞軸索の伸展など、微小管の働きを介して行われます。微小管はまた、有糸分裂/減数分裂中の紡錘体形成においても重要な役割を果たし、分裂中期の染色体の整列も担っています。微小管は、中心体の重要な構成分子である中心小体の9+2構造を形成し、これはMicrotubule-organizing center (MTOC) として働き、細胞極性においても役割を果たしています。有糸分裂におけるその役割から、微小管はがん化学療法の標的とされてきました。微小管は常に動的不安定な状態にあり、プラス端での微小管の重合と、マイナス端での脱重合が競合しています。このプロセスはStathmin、Diap1/2、Tau、低分子量GTPaseのRhoファミリーなど、複数のシグナル伝達分子によって制御されています。
マイクロフィラメントは、細胞骨格の主要な構成分子であり、Actinの繊維状重合体であるF-Actinから構成されます。マイクロフィラメントは細胞の形態変化、移動、増殖および生存において重要です。アクチン細胞骨格の制御は、Gタンパク質共役型受容体 (GPCR)、インテグリン、受容体チロシンキナーゼ (RTK) や、Semaphorin 1a受容体のPlexin Aのような他の数多くの特異的な受容体を介するシグナル伝達から開始されます。これらの受容体によって、非常に多くのシグナル伝達カスケードが開始され、これには低分子GTPaseのRhoファミリー (Rho、Rac、Cdc42) とそれらの活性化因子、グアニンヌクレオチド交換因子 (GEF) とその下流にあるRho-キナーゼ/ROCKおよびp21 activated kinase (PAK) を含むプロテインキナーゼエフェクターが含まれます。また、Cortactin、Diap1/2、WAVE、WASPなどの複数のアクチン調節タンパク質へのGTPaseの直接的な結合も、これらの受容体を介して起こります。これらのカスケードは、Cofilin、Arp2/3複合体、Ena/VASP、Profilin、Gelsolinのようなアクチン相互作用性の調節タンパク質など、アクチン細胞骨格の挙動や構成を直接的に制御するタンパク質に集中しています。
中間径フィラメントの主要なタイプは、その細胞特異的な発現によって明確に区別されます。それには、サイトケラチン (上皮細胞)、Glial Fibrillary Acidic Protein (GFAP:グリア細胞)、Desmin (骨格筋細胞、内臓筋細胞、および特定の血管平滑筋細胞)、Vimentin (間充織の細胞)、および神経フィラメント (神経細胞) が含まれます。このグループのメンバーは、球状のアミノ末端のヘッドドメイン、中央のα−Tubulinロッドドメイン、およびカルボキシ末端のテールを含みます。中間径フィラメントは細胞の構造を支え、細胞小器官やモーター分子の固定点として働き、内因性ストレスや環境ストレスから細胞を防御するストレスタンパク質としての機能も有しています。中間径フィラメントの制御は、リン酸化を含む複数のメカニズムを介して行われます。そして、それは他の中間径フィラメントと相互作用するタンパク質との集合状態や、集合活性に影響を及ぼします。
参考文献:
- Goldman RD, Grin B, Mendez MG, Kuczmarski ER (2008) Intermediate filaments: versatile building blocks of cell structure. Curr. Opin. Cell Biol. 20(1), 28–34.
- Toivola DM, Strnad P, Habtezion A, Omary MB (2010) Intermediate filaments take the heat as stress proteins. Trends Cell Biol. 20(2), 79–91.
- Ridley AJ (2011) Life at the leading edge. Cell 145(7), 1012–22.
- Rottner K, Stradal TE (2011) Actin dynamics and turnover in cell motility. Curr. Opin. Cell Biol. 23(5), 569–78.
- Windoffer R, Beil M, Magin TM, Leube RE (2011) Cytoskeleton in motion: the dynamics of keratin intermediate filaments in epithelia. J. Cell Biol. 194(5), 669–78.
- Sandquist JC, Kita AM, Bement WM (2011) And the dead shall rise: actin and myosin return to the spindle. Dev. Cell 21(3), 410–9.
- Field CM, La¡rt P (2011) Bulk cytoplasmic actin and its functions in meiosis and mitosis. Curr. Biol. 21(19), R825–30.
- Aylett CH, Lawe J, Amos LA (2011) New insights into the mechanisms of cytomotive actin and tubulin filaments. Int Rev Cell Mol Biol 292, 1–71.
- Bornens M (2012) The centrosome in cells and organisms. Science 335(6067), 422–6.