

抗体の検証における戦略:バイナリー戦略
抗体検証にバイナリー戦略を採用するCSTアプリケーション
バイナリーアプローチは、抗体の特異性を評価する最良の方法の1つです。生物学的に関連するポジティブ発現系とネガティブ発現系で抗体を試験することにより、サンプルに存在する他の生体分子と交差反応することなく、未変性状態における標的抗原の認識能力を確認できます。バイナリーモデルでは、標的タンパク質の発現がポジティブ (高い) あるいはネガティブ (低い) であることが既知または予測可能な細胞や組織における内在性の標的、遺伝的ノックアウト、標的タンパク質の発現や修飾を誘導あるいは阻害する処理を利用します。
バイナリー試験を効果的に行うためには、遺伝子シーケンシングによるノックアウトの確認やプロテオミクスプロファイリングによる発現レベルの確認など、直交的な手法を用いてデータを常に確認する必要があります。抗体の性能に最大限の信頼を持たせるため、その他の検証戦略も採用する必要があります。また、抗体のバイナリー検証に使用するモデルはそれぞれ、抗体の使用を予定しているアプリケーションごとに試験する必要があります。ある抗体がウェスタンブロットで特異性を示しても、同じ抗体が免疫組織化学染色 (IHC) でも同様に特異性を示すとは限りません。
内在性コントロール
目的の標的を内在的に発現する細胞や組織は、最も一般的なポジティブコントロールです。標的を発現していない似たサンプルを比較に用いることにより、抗体による抗原の特異的な認識の有無を迅速に確認できます。
この戦略によって意義ある知見を確実に得るには、適切なポジティブコントロールあるいはネガティブコントロールとなるサンプルを選択することが重要です。そのために、ゲノムデータベースやトランスクリプトームデータベース、プロテオミクスデータベースを利用して大量の文献を確認しています。あるいは、試験する抗体によって得られた免疫染色を、同じ標的の異なる抗原に対する、すでに完全に検証された抗体の免疫染色と比較します。
図1および図2は、内在性コントロールを用いた抗体検証の例です。抗体検証に使用するモデルは、抗体の使用を予定しているアプリケーションごとに試験する必要があることが明確に分かります。図1は、Jurkat細胞とRL細胞をHelios (E4L5U) Rabbit Monoclonal Antibodyを用いてウェスタンブロッティングで解析しています。Heliosの発現はJurkat細胞でポジティブ、RL細胞でネガティブとなっています。一方、図2は、同じ細胞モデルを用いてこの抗体を免疫組織化学染色 (IHC) で解析しています。ローディングコントロールを用いてサンプルの品質に問題が無いことを確認しており、ウェスタンブロットとIHCのデータに相違はありません。
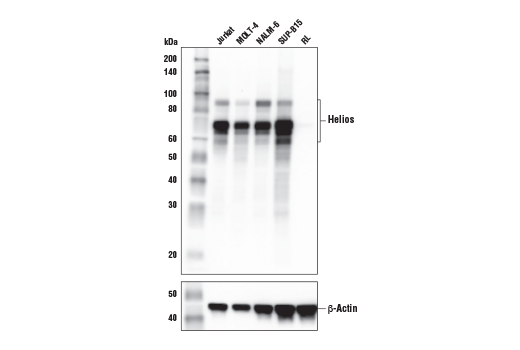
図1 様々な細胞株からの抽出物を、Helios (E4L5U) Rabbit Monoclonal Antibody (上) およびbeta-Actin (D6A8) Rabbit Monoclonal Antibody #8457 (下) を用いてウェスタンブロッティングで解析しました。予想通り、Heliosタンパク質はRL細胞で発現がみられません。

図2 パラフィン包埋Jurkat細胞ペレット (左、ポジティブ) またはRL細胞ペレット (右、ネガティブ) を、Helios (E4L5U) Rabbit Monoclonal Antibodyを用いて免疫組織化学染色で解析しました。
遺伝子ノックアウト
普遍的に発現している一部のタンパク質を検証する場合や、発現をサポートするデータが矛盾しているまたは信頼性を欠く場合など、内在性のバイナリーモデルが常に利用できるとは限りません。このような場合には、マウスのノックアウトモデルが抗体の特異性のバイナリー評価に広く用いられます。通常、共同研究者から提供されるマウスのノックアウトサンプルは、野生型およびノックアウト型の細胞や組織で構成されています。これらはそれぞれ、標的抗原の未変性状態における発現と未発現を示すサンプルのため、利用が可能であれば理想的な試験パラダイムとなります。
遺伝的ノックアウトを実現するもう1つの方法として、CRISPRを介した遺伝子破壊を利用する方法があります。これは近年、直感的で柔軟性の高いシステムに簡略化され、その有用性が高まってきています。このアプローチの利点は、同じ細胞株でポジティブおよびネガティブの両方の発現モデルが得られることです。ただし、遺伝子の破壊は常に完全であるとは限らず、遺伝子の部分的な欠失や断片化が起こり、これに由来するシグナルが得られることがあるので注意が必要です。標的のノックダウンは、抗体に依存しない直交的な方法で確認することが重要です。また、細胞のCRISPRノックアウトは、細胞ベースのアッセイで抗体の性能を検証する場合は非常に有効ですが、様々な細胞タイプが混在する免疫組織化学染色などの組織ベースのアッセイには必ずしも有効では無いことに注意が必要です。
CRISPR以外にも、合成2本鎖RNA (siRNA [small/short interfering RNA]) を用いて標的タンパク質をコードする遺伝子の発現を一過性に抑制するノックダウンも一般的な方法です。さらに別の選択肢として、DNAベクターに組み込んだshRNA (short/small hairpin RNA) をトランスフェクションやウイルス感染により細胞に導入する方法もあります。ベクターのセレクションマーカーを利用して、トランスフェクションまたはウイルス感染した細胞を選選別し、宿主ゲノムにshRNAが安定に組み込まれ、長期にわたって遺伝子がサイレンシングされた細胞を得ることもできます。
この場合も、抗体検証に使用する遺伝子ノックアウトモデルは、抗体の使用を予定しているアプリケーションごとに試験することが非常に重要です。図3および図4では、野生型マウスおよびRKIPノックアウトマウスの胚性線維芽細胞 (MEF) をRKIP (D42F3) Rabbit Monoclonal Antibodyを用いてウェスタンブロッティングと免疫組織化学染色で解析しました。図5および図6では、野生型およびThymidine Kinase 1ノックアウトHCT 116細胞をThymidine Kinase 1 (E2H7Z) Rabbit Monoclonal Antibodyを用いてウェスタンブロッティングと免疫細胞化学染色で解析しました。IHCとウェスタンブロットのデータに相違はありません。
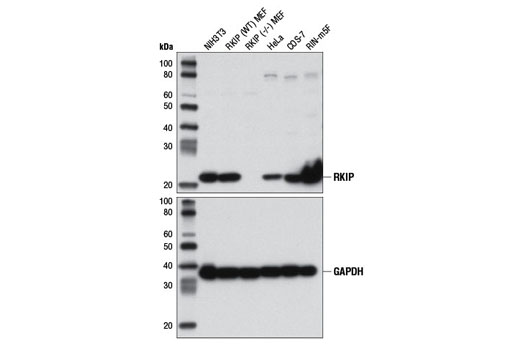
図3 RKIP野生型 (WT) およびRKIPノックアウト (-/-) MEF細胞を含む様々な細胞株からの抽出物を、RKIP (D42F3) Rabbit Monoclonal Antibody (上) あるいはGAPDH (D16H11) Rabbit Monoclonal Antibody #5174 (下) を用いてウェスタンブロッティングで解析しました。RKIP MEF細胞は、シカゴ大学のMarsha Rosner博士のご厚意により提供いただきました。
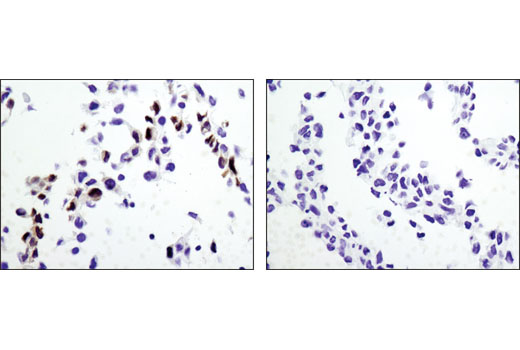
図4 パラフィン包埋した野生型MEF (左) またはRKIPノックアウトMEF (右) の細胞ペレットを、RKIP (D42F3) Rabbit Monoclonal Antibodyを用いて免疫組織化学染色で解析しました。MEFは、シカゴ大学のMarsha Rosner博士のご厚意により提供いただきました。
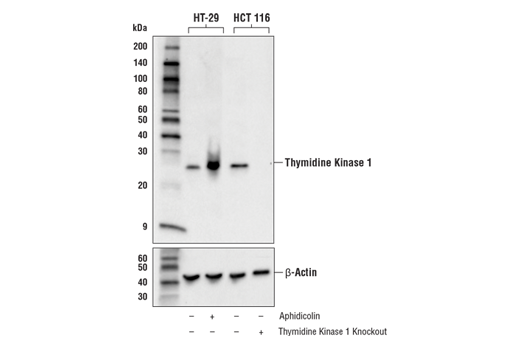
図5 10 μg/mLのAphidicolinで24時間処理したHT-29細胞 (+)、未処理のHT-29細胞 (-)、野生型HCT 116細胞 (-)、Thymidine Kinase 1ノックアウトHCT 116細胞 (+) からの抽出物を、Thymidine Kinase 1 (E2H7Z) Rabbit Monoclonal Antibody (上) またはbeta-Actin (D6A8) Rabbit Monoclonal Antibody #8457 (下) を用いてウェスタンブロッティングで解析しました。
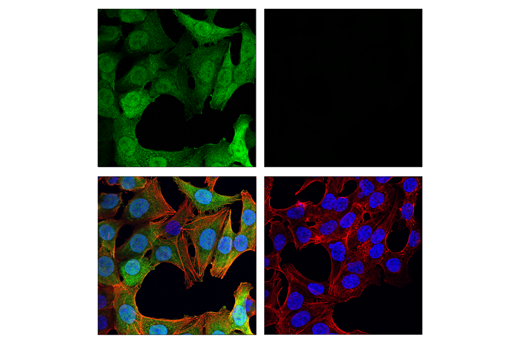
図6 野生型 (左、ポジティブ) またはThymidine Kinase 1ノックアウト (右、ネガティブ) のHCT 116細胞を、Thymidine Kinase 1 (E2H7Z) Rabbit Monoclonal Antibody (緑) を用いて免疫蛍光染色し、共焦点顕微鏡で観察しました。アクチンフィラメントはDyLight 554 Phalloidin #13054 (赤) で染色しています。サンプルは、ProLong® Gold Antifade Reagent with DAPI #8961 (青) で封入しました。
標的抗原の修飾
バイナリーモデルでは、他にもタンパク質標的の発現および局在、翻訳後修飾 (PTM) の誘導あるいは阻害 (アゴニストあるいはアンタゴニストを用いた処理) を使用する方法があります。これらの処理で、ほぼ完全なポジティブシグナルとネガティブシグナルが得られる場合は、バイナリー検証に利用できます。標的抗原の修飾は、タンパク質のリン酸化やアセチル化など、翻訳後修飾された標的に対する抗体を検証するのに適したアプローチです。
標的抗原の修飾に依存する検証法の場合も、他の検証法と同様に、適切なローディングコントロール、発現コントロールは非常に重要です。これらにより、処理の有効性や、評価対象のサンプルが質、量ともに同等を確認できます。処理が、できる限り標的に特異的であることも重要です。広範な標的から非特異的に翻訳後修飾を除去する一般的なホスファターゼや脱アセチル化酵素で処理を行う前に、例えば、タンパク質や経路に特異的なアゴニストや阻害剤について検討すべきです。
図7および図8では、フィブロネクチンでコーティングしたプレート表面への細胞接着を利用してPaxillinのチロシン残基のリン酸化を誘導したHeLa細胞を用いて、Phospho-Paxillin (Tyr118) 抗体の検証を行いました。
ポジティブコントロールとネガティブコントロールの組み合わせ
免疫組織化学染色で最も頻繁に使用されるその他のバイナリー戦略に、同じ切片にポジティブ細胞とネガティブ細胞の両方を含む組織サンプルを用いて抗体を検証する方法があります。この例を図9および図10に示しています。観察された染色パターンは、標的の既知の発現プロファイルに一致しています (すなわち、発現の無い細胞は染色されず、適切な細胞がポジティブ染色されています)。
バイナリーモデルは抗体試薬の検証に有効な手段ですが、必ずしも材料を容易に入手できたり、速やかに準備ができる訳ではありません。バイナリー試験のデータをサポートするため、また抗体の性能を評価する代替の手段として、追加の検証方法を常に利用する必要があります。
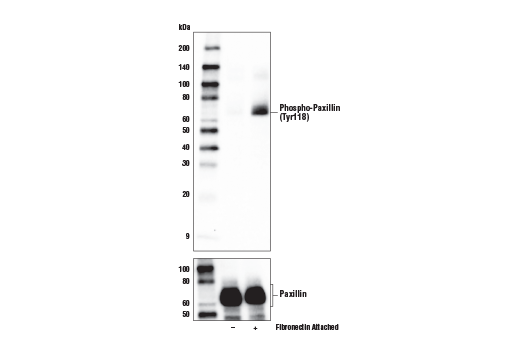
図7懸濁した未処理のHeLa細胞 (-) または10 μg/mLフィブロネクチンでコーティングしたプレート表面へ1時間接着させたHeLa細胞 (+) からの抽出物を、Phospho-Paxillin (Tyr118) (E9U9F) Rabbit Monoclonal Antibody (上) あるいはPaxillin (D9G12) Rabbit Monoclonal Antibody #12065 (下) を用いてウェスタンブロッティングで解析しました。
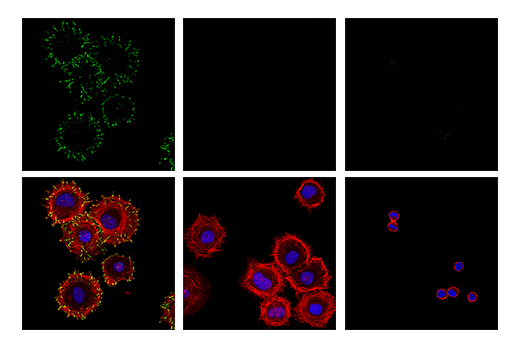
図8 HeLa細胞は0.04%トリプシンおよび0.03% EDTAで剥離し、無血清培地で1時間懸濁させ、細胞を懸濁させたまま (右) または固定の前にフィブロネクチンでコーティングしたスライド上に播種し、さらに1時間接着させました (左)。また、リン酸化の特異性を確認するため、接着させた細胞をλ-ホスファターゼで後処理しました (中央)。
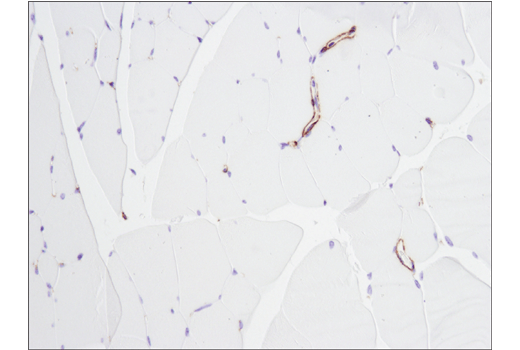
図9 パラフィン包埋マウス骨格筋組織を、alpha-Smooth Muscle Actin (D4K9N) Rabbit Monoclonal Antibodyを用いて免疫組織化学染色で解析しました。予想通り、血管平滑筋細胞が染色され、平滑筋アクチンの発現の無い筋細胞は染色されていません。
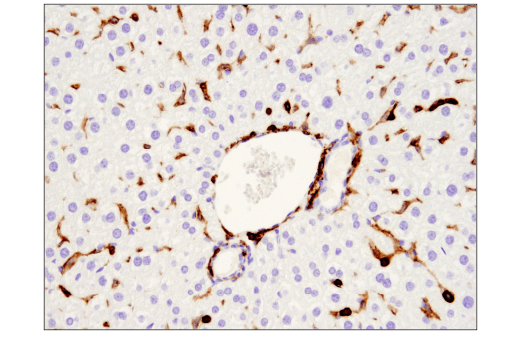
図10 パラフィン包埋したマウス肝臓組織を、CD45 (D3F8Q) Rabbit Monoclonal Antibodyを用いて免疫組織化学染色で解析しました。予想通り、CD45ネガティブ肝細胞は染色されていません。