View in English?
View in English?
View in English?

| Cat. # | Size | Qty. | Price | Inventory |
|---|---|---|---|---|
| 9653T | 1 Kit (9 x 20 microliters) |
|
| Product Includes | Quantity | Applications | Reactivity | MW(kDa) | Isotype |
|---|---|---|---|---|---|
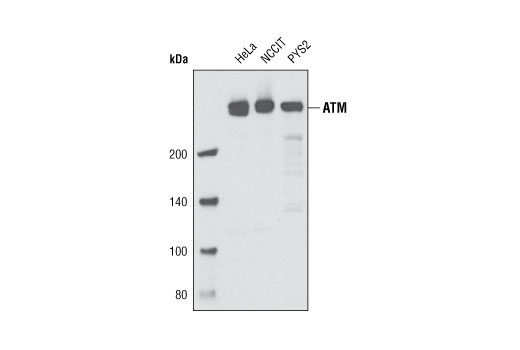
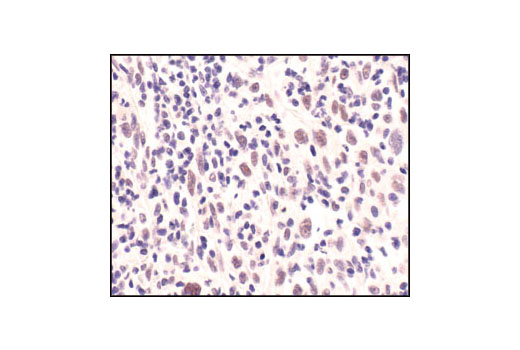
| Phospho-ATM (Ser1981) (D6H9) Rabbit mAb 5883 | 20 µl |
|
H | 350 | Rabbit IgG |
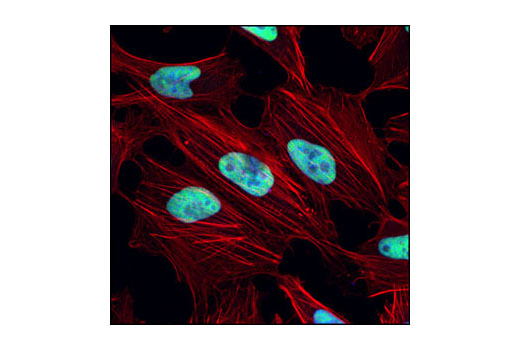
| ATM (D2E2) Rabbit mAb 2873 | 20 µl |
|
H M | 350 | Rabbit IgG |
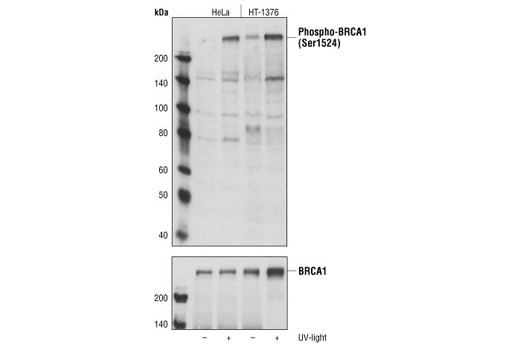
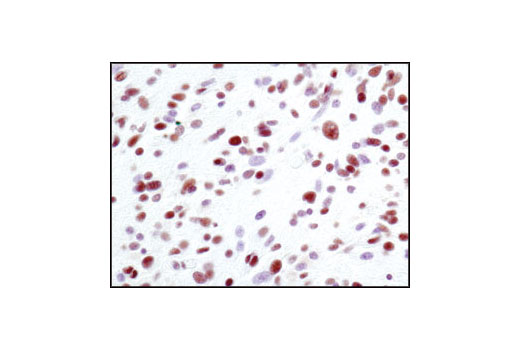
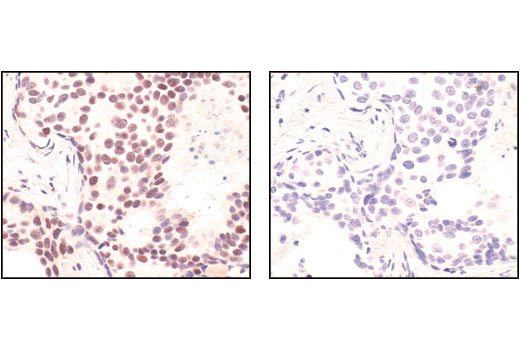
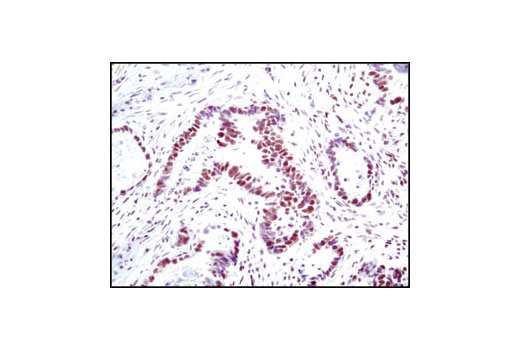
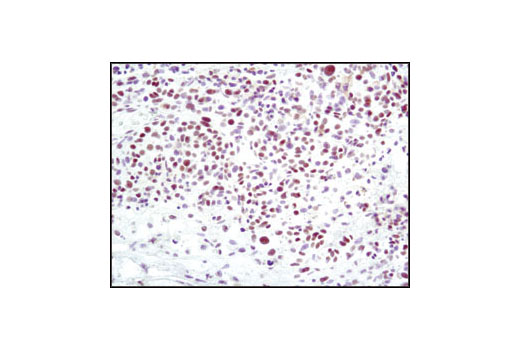
| Phospho-BRCA1 (Ser1524) Antibody 9009 | 20 µl |
|
H | 220 | Rabbit |
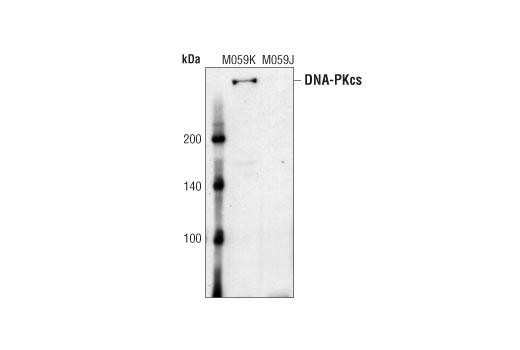
| DNA-PKcs Antibody 4602 | 20 µl |
|
H | 450 | Rabbit |
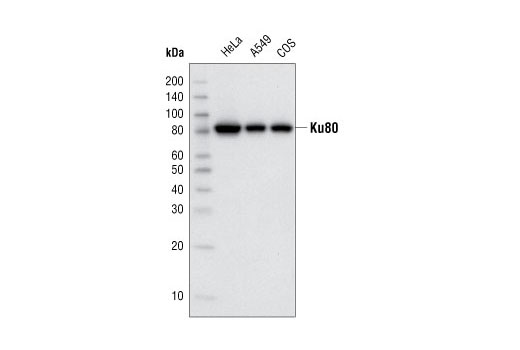
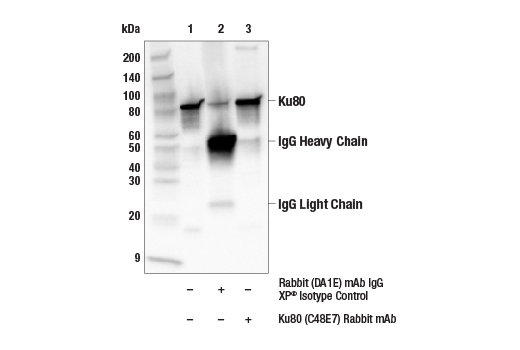
| Ku80 (C48E7) Rabbit mAb 2180 | 20 µl |
|
H Mk | 86 | Rabbit IgG |
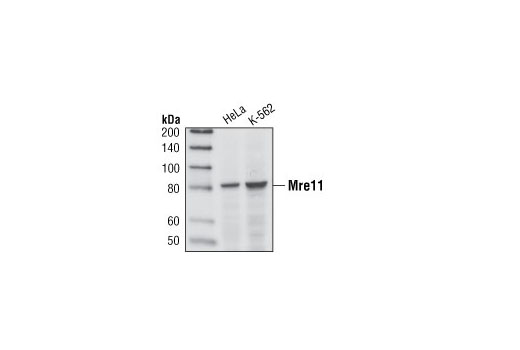
| Mre11 (31H4) Rabbit mAb 4847 | 20 µl |
|
H | 81 | Rabbit IgG |
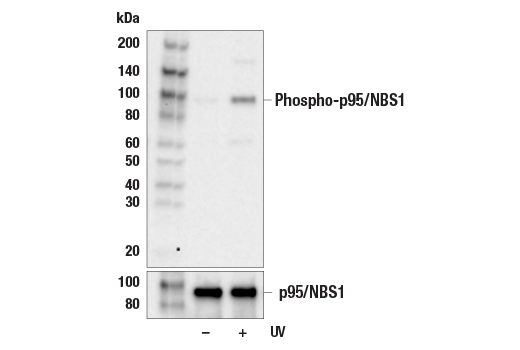
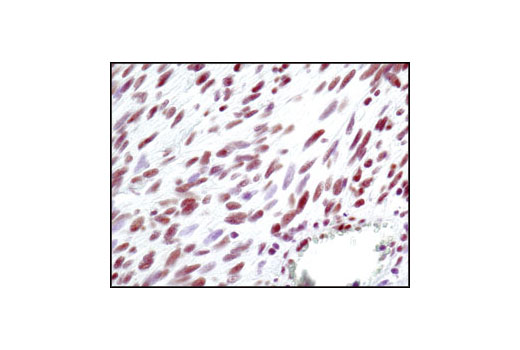
| Phospho-p95/NBS1 (Ser343) Antibody 3001 | 20 µl |
|
H | 95 | Rabbit |
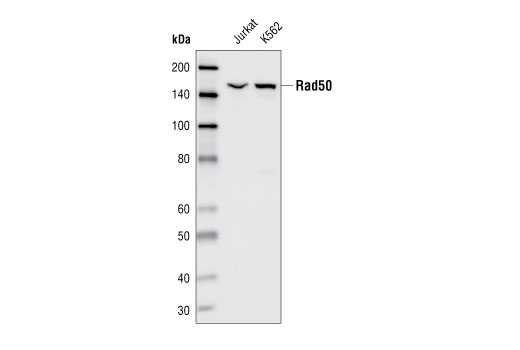
| Rad50 Antibody 3427 | 20 µl |
|
H Mk | 153 | Rabbit |
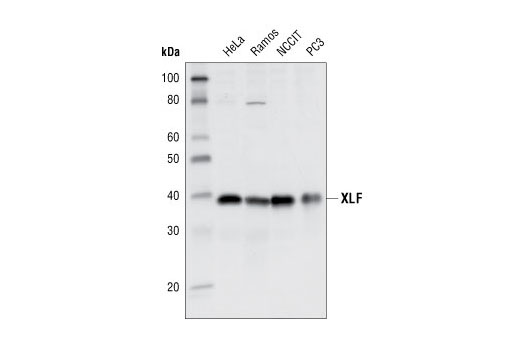
| XLF Antibody 2854 | 20 µl |
|
H | 39 | Rabbit |
| Anti-rabbit IgG, HRP-linked Antibody 7074 | 100 µl |
|
Goat |
Product Information
Monoclonal antibodies are produced by immunizing animals with synthetic peptides corresponding to residues surrounding Lys496 of human Mre11A and the carboxy terminus of human Ku80. Activation state monoclonal antibody is produced by immunizing animals with a synthetic phosphopeptide corresponding to residues around Ser1981 of human ATM. ATM (D2E2) Rabbit mAb is produced by immunizing animals with recombinant human ATM. Polyclonal antibodies are produced by immunizing animals with synthetic peptides corresponding to the carboxy-terminus of human DNA-PKcs, the amino terminus of human Rad50, and the carboxy terminus of human XLF. Activation state polyclonal antibodies are produced by immunizing animals with synthetic phosphopeptides corresponding to residues surrounding Ser1524 of human BRCA1 and surrounding Ser343 of human p95/NBS1. Polyclonal antibodies are purified by protein A and peptide affinity chromatography.
Double strand DNA breaks (DSB) in mammalian cells can be repaired by the related mechanisms of non-homologous end-joining (NHEJ) and homologous recombination (HR). A DNA-dependent protein kinase composed of DNA-binding subunits Ku70 and Ku86 and the DNA-PKcs catalytic subunit mediates NHEJ repair. The Ku heterodimer binds free DNA ends and recruits DNA-PKcs to the break (1). DNA-PKcs signals areas of DNA damage and recruits additional proteins, such as the Artemis exo- and endonuclease that processes and primes the damaged sequence (2,3). Following replacement DNA synthesis, a ligase complex composed of DNA ligase IV and XRCC4 joins the repaired ends. XRCC4-like factor (XLF) is an essential ligase-associated repair factor that promotes gap-filling during NHEJ (4). Homologous recombination utilizes aligned homologous sequences as a repair template. The MRN complex, composed of Mre11, Rad50, and nibrin (p95/NBS1), plays a critical role in sensing, processing and repairing breaks (5). MRN interacts with BRCA1 and CtIP to facilitate 5’ resection of DSB DNA to generate 3’ ssDNA ends necessary for repair (6). DNA-binding protein Mre11 exhibits exonuclease and endonuclease activity and is largely responsible for ssDNA end processing (7). Interaction between the MRN complex and ATM kinase promotes association between the kinase and its substrates and likely leads to ATM activation (8). ATM acts a central controller of the cell cycle checkpoint by phosphorylating multiple targets, including c-Abl, BRCA1 and p95/NSB1. Activated c-Abl phosphorylates Rad52, which promotes Rad51 binding to ssDNA and subsequent annealing of ssDNA (7).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.
View in English?