

細胞死の誘導
アポトーシス、マイトファジー、ネクロプトーシス、オートファジーなどの細胞死パスウェイの変異は、神経細胞死や神経変性疾患の進行に寄与します。アポトーシス促進性および抑制性シグナル伝達の異常、ミトコンドリアの機能不全、オートファジーや小胞体ストレス応答の誤制御、ストレスや炎症によるネクロソームの活性化は、神経細胞死や疾患を引き起こすメカニズムのごく一部です。これらのパスウェイの多くは非神経細胞での理解が進んでいますが、ニューロンにおけるその活性化メカニズムや調節不全については謎のまま独自の課題が残されています。
以下の標的から始めませんか?
切断型PARP (Asp214)
PARPは通常、酸化ストレスに応答したDNA修復パスウェイの重要な制御因子として機能します。Caspase-3によってAsp214とGly215の間が切断されると、切断されたN末端側の断片は、DNA修復酵素を阻害してニューロンをアポトーシスに向け、アポトーシス細胞の特性を示すようになります。
製品
Cleaved PARP (ASP214) (D64E10) XP® Rabbit mAb #5625 – IF
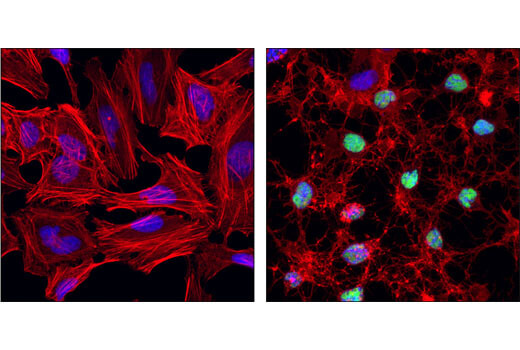
Cleaved PARP (ASP214) (D64E10) XP® Rabbit mAb #5625: 未処理コントロール (左) あるいはStaurosporine #9953で処理 (右) したHeLa細胞を、Cleaved PARP (Asp214) (D64E10) XP® Rabbit mAb (緑) を用いて免疫蛍光染色し、共焦点顕微鏡で観察しました。アクチンフィラメントをDY-554 phalloidinで染色しています。DRAQ5 #4084 (蛍光DNA染料) は青の疑似カラーで示しました。
PINK1
PINK1 (PTEN-induced kinase 1) は、ミトコンドリアのセリン/スレオニンプロテインキナーゼで、ストレス誘導性のミトコンドリア機能障害から細胞を保護します。これはひどく損傷したミトコンドリアの外膜に蓄積し、PARKINをリクルートしてオートファジーによる分解を誘導します。PINK1の変異は、常染色体劣性早発性パーキンソン病と関連しています。
製品
PINK1 (D8G3) Rabbit mAb #6946 – W, IP
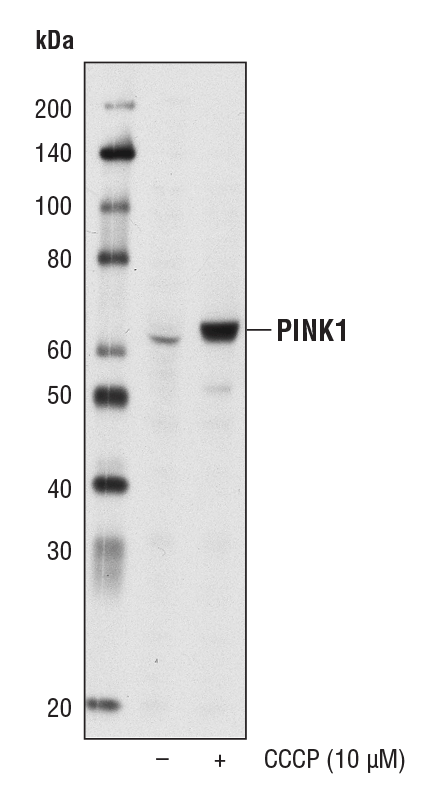
PINK1 (D8G3) Rabbit mAb #6946: 未処理コントロール (-) あるいはCCCP 10 μMで24時間処理 (+) したHeLa細胞の抽出物を、PINK1 (D8G3) Rabbit mAbを用いてウェスタンブロッティングにより解析しました。
SQSTM1/p62
Sequestosome 1 (SQSTM1、p62) はオートファゴソームのカーゴタンパク質で、タンパク質凝集体に結合して選択的オートファジーによる分解に導きます。SQSTM1/p62の変異は、それぞれパーキンソン病、ハンチントン病、アルツハイマー病の進行を促進させる、α-Synuclein、Huntingtin、Tauタンパク質およびβ-Amyloidの細胞内凝集体を増加させます。
製品
SQSTM1/p62 (D10E10) Rabbit mAb (IF Preferred) #7695 – IP, IF-IC
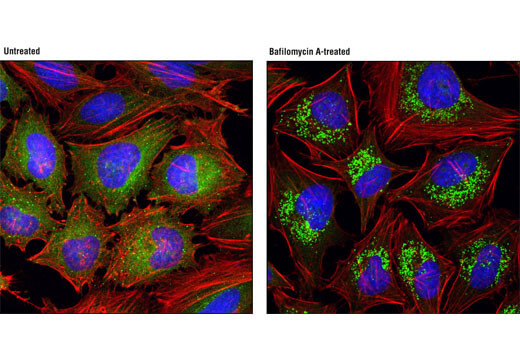
SQSTM1/p62 (D10E10) Rabbit mAb (IF Preferred) #7695: 未処理コントロール (左) あるいはBafilomycin A 100 nMで18時間処理 (右) したHeLa細胞を、SQSTM1/p62 (D10E10) Rabbit mAb (IF Preferred) (緑) を用いて免疫蛍光染色し、共焦点顕微鏡で観察しました。アクチンフィラメントをDY-554 phalloidinで染色しています。DRAQ5 #4084 (蛍光DNA染料) は青の疑似カラーで示しました。
LC3A/B
LC3A/Bはオートファゴソームの形成と成熟に重要な役割を果たし、オートファゴソームのカーゴを選択的にリクルートするアダプタータンパク質としても機能します。TREM2変異をもつアルツハイマー病患者の組織ではLC3陽性ミクログリアの増加が見られ、TREM2依存的なオートファジーの阻害がアルツハイマー病の病因に寄与する可能性があります。
製品
LC3A/B (D3U4C) XP® Rabbit mAb #12741 – W, IHC-P, IF-IC, F
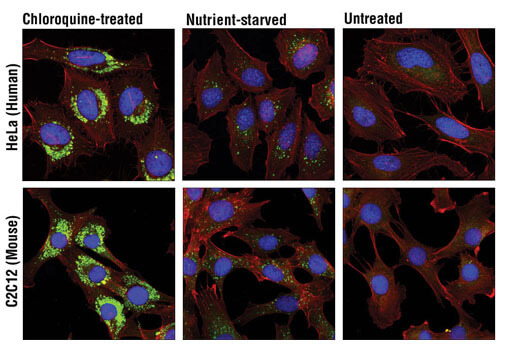
LC3A/B (D3U4C) XP® Rabbit mAb #12741: Chloroquine 50 µMで一晩処理 (左)、EBSSで栄養飢餓を3時間誘導 (中央)、未処理コントロール (右) のHeLa細胞 (上段) とC2C12細胞 (下段) を、LC3A/B (D3U4C) XP® Rabbit mAb (緑) およびβ-Actin (13E5) Rabbit mAb (Alexa Fluor® 555 Conjugate) #8046 (赤) を用いて免疫蛍光染色し、共焦点顕微鏡で観察しました。DRAQ5 #4084 (蛍光DNA色素) は、青の疑似カラーで示しています。
Phospho-RIP3 (Ser227)
ヒトリン酸化RIP3 (Ser227) は、MLKL1をリン酸化してTNF誘導性ネクロトーシスを引き起こします。この形態のプログラムされた細胞死は、多発性硬化症と筋萎縮性側索硬化症で報告されています。
製品
Phospho-RIP3 (Ser227) (D6W2T) Rabbit mAb #93654 – W, IF-IC
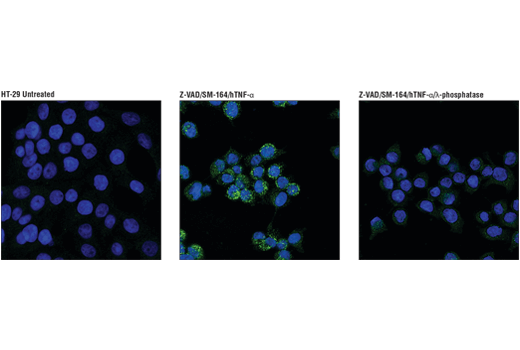
Phospho-RIP3 (Ser227) (D6W2T) Rabbit mAb #93654:未処理コントロール (左)、Z-VAD 20 µMで30分間前処理した後SM-164 100 nMおよびHuman Tumor Necrosis Factor-α (hTNF-α) #8902 20 ng/mLで6時間処理 (中央)、Z-VADで前処理した後SM-164、hTNF-αおよびプロセシング済みホスファターゼで処理 (右) したHT-29細胞を、Phospho-RIPK3 (Ser227) (D6W2T) Rabbit mAb (緑) を用いて免疫蛍光染色し、共焦点顕微鏡で観察しました。アクチンフィラメントは、DyLight554 Phalloidin #13054 (赤) で染色しました。DRAQ5 #4084 (蛍光DNA染料) は青の疑似カラーで示しました。