免疫学の概要
免疫系
免疫系は組織、細胞、分子から成り立っており、その主要機能は病原体や形質転換した細胞を検出して排除することです。
自然免疫応答と適応免疫応答
免疫系には大きく分けて (1) 自然免疫系、(2) 適応免疫系の2つがあります。自然免疫系は病原体の検出と生体防御の第一線であり、生殖系列にコードされたパターン認識受容体を介した反応で、記憶の形成は起こらず迅速に応答します (数分から数時間以内)。適応免疫は発生段階で複数の遺伝子再編成を経た抗原特異的な受容体を介した反応で、時間をかけて起こる応答です (数日以上)。この応答では初めて遭遇した病原体に対して学習 (免疫記憶) が起こり、将来同様の有害物質に遭遇した場合にこれと戦う備えをします。どちらのタイプの免疫応答も細胞外液中の巨大分子や特定の免疫細胞の活性化によって引き起こされます。これらの応答はそれぞれ液性免疫、細胞性免疫と呼ばれています。
細胞性免疫応答では、免疫細胞は主に自然免疫担当細胞と適応免疫担当細胞に分類されます。自然免疫系を担当する主な細胞には好中球、マクロファージ、ナチュラルキラー細胞、好塩基球など、侵入してくる危険な微生物を標的とする食細胞があります。適応免疫系を担当する細胞はT細胞とB細胞です。ナチュラルキラー (NK) T細胞は自然免疫と獲得免疫の細胞の特色を併せもっています。また、ほとんどの細胞 (免疫系に特化していない他のタイプの細胞) は細胞内自然免疫機能をもち、RIG-1、STING、NLRファミリーのメンバーなどのエフェクター分子を介して細胞質受容体の形成やシグナル伝達を行います。後記のように、自然免疫と獲得免疫のどちらの系の細胞も造血幹細胞に由来する異なる細胞系列 (骨髄系とリンパ系) から分化します。
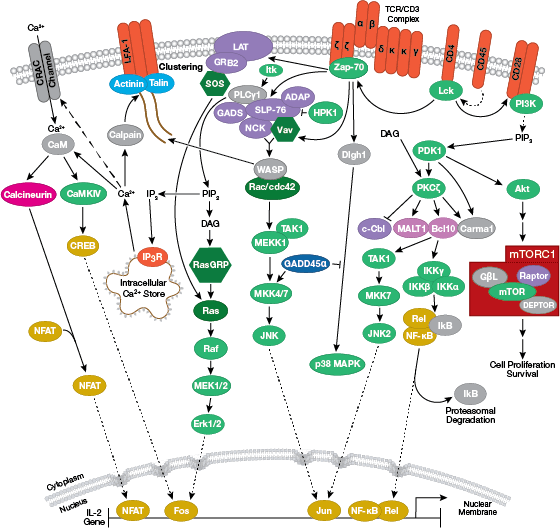
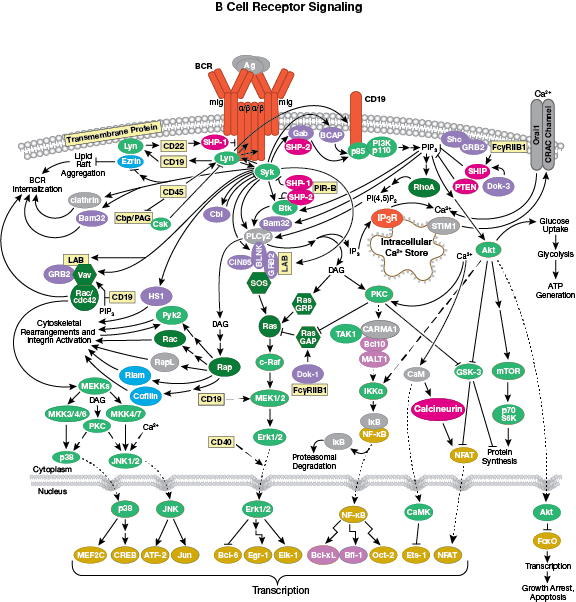
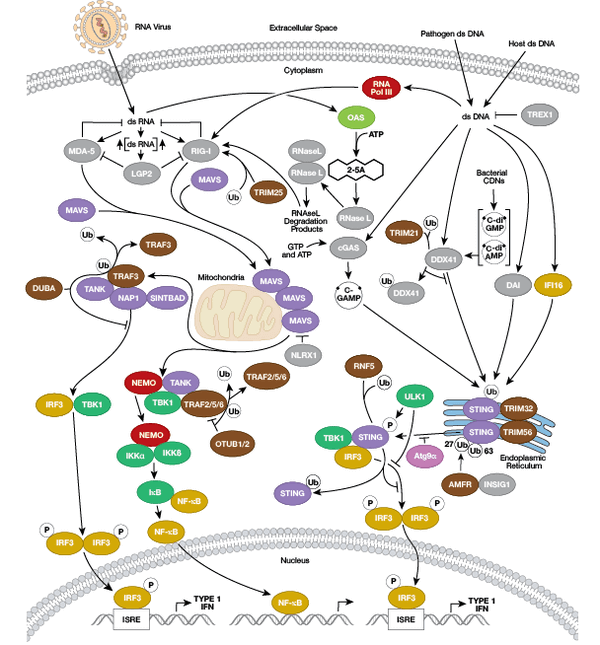
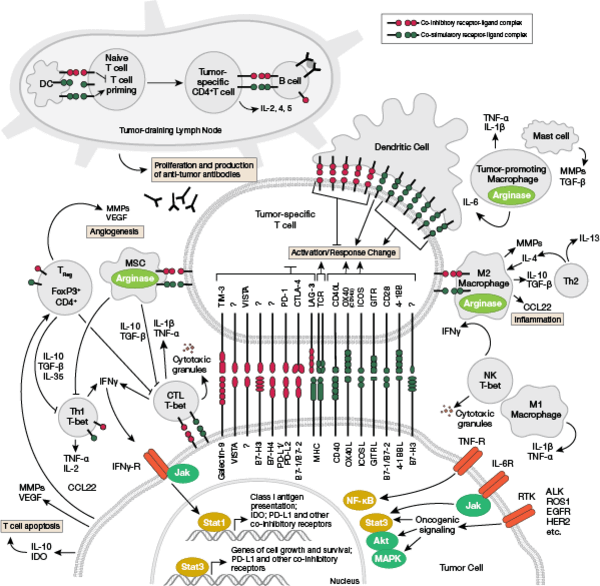
免疫系の構成因子
成熟免疫細胞は造血幹細胞から分化します。これらは多能性をもつ未分化前駆細胞で、骨髄や末梢血、胎盤などにみられます。造血幹細胞は骨髄系共通前駆細胞またはリンパ系共通前駆細胞に分化することができます。
リンパ系細胞
リンパ系共通前駆細胞は大きく分けてT細胞、B細胞、ナチュラルキラー (NK) 細胞、NK-T細胞の4つの集団に分化することができ、これらは細胞表面受容体の発現によって識別することができます。
T細胞
T細胞は細胞性免疫で重要な役割を果たします。これらの細胞はT細胞シグナル伝達鎖CD3の発現によって特定することができます。
T細胞は骨髄の造血幹細胞に由来するリンパ系前駆細胞から分化します。リンパ系前駆細胞はT細胞への分化が決定づけられると骨髄から胸腺 (thymus) に移動します (このためT細胞と名付けられました)。胸腺はT細胞が様々なサブタイプに分化するのに適した微小環境を提供します。これらのサブタイプは細胞表面に発現する特異的な受容体によって区別することができます。成熟の早期段階において、T細胞 (または胸腺細胞) はCD4、CD8のどちらの受容体も発現しておらず、このためダブルネガティブ (DN) 細胞と呼ばれます。DN細胞はその後、T細胞受容体 (TCR) α鎖とβ鎖をコードする遺伝子の遺伝子再編成と変異を受けます。これによってT細胞分化の下流経路が開始され、最終的に固有の抗原に特異的な親和性をもつT細胞受容体が出来上がります。
胸腺細胞はこの後、正の選択と負の選択を受け、自己抗原には反応せずMHCを介して細胞表面にペプチドとして提示された「外来」抗原のみを認識するものが残ります。この2つのテストで選択された細胞のみがCD4とCD8を発現するようになり、ダブルポジティブ細胞となります。成熟の後期段階においてT細胞はCD4かCD8のどちらかを発現するようになり (他方の発現がなくなり)、シングルポジティブ細胞となります。これらCD4またはCD8シングルポジティブT細胞は血流に放出されます。放出されたT細胞は、この時点で「ナイーブ」状態で、特定のT細胞サブタイプへの分化の決定づけはなされていません。
成熟過程で自己のMHCに中程度の親和性を示した胸腺細胞は、有害な「自己」抗原を認識する能力により制御性T細胞 (Treg) になるように選択される場合があります。血中や末梢組織のT細胞も局所的に誘導されてTregになる場合があります。Tregの主な役割は自己抗原に対する寛容性を維持することと、T-エフェクター細胞の機能と増殖を制限することです。
血中や末梢組織に到達したナイーブT細胞はヘルパーT細胞への分化誘導を受けることができます。マクロファージ、樹状細胞、B細胞などの抗原提示細胞 (APC) がT細胞に抗原を提示し、その受容体が抗原に親和性をもっていた場合にT細胞は抗原を認識します。通常、CD4+ T細胞がMHCクラスIIに結合した抗原ペプチドの認識に適しています。ペプチド-MHC複合体が認識されるとT細胞は一連の細胞内シグナル伝達経路を活性化させ、抗原の認識が確認されます。ここで初めてヘルパーT細胞が増殖し、様々な細菌や寄生生物など有害な抗原に特異的な細胞のプールが拡張されます。
抗原を認識して活性化されたヘルパーT細胞は様々なサイトカインを分泌し、他の免疫細胞に有害分子への適切な対処法を伝達します。ヘルパーT細胞にはTh1、Th2、Th17などのいくつかのサブセットがあります。これらのサブセットは異なるサイトカインを産生、分泌して、病原の種類によって免疫応答を適切に調整する助けとなります。例えば、Th1細胞はエフェクターサイトカインIFN-γを介してマクロファージにシグナルを伝達して有害物質の消化や破壊を促進するのに対し、Th2細胞はIL-4、IL-5、IL-9、IL-10、IL-13を組み合せて分泌して好塩基球、好酸球、その他の細胞にシグナルを伝達して感染に対処します。サイトカインの種類や量によって感染や有害な細胞への適切な応答が決まります。T細胞は細胞傷害性T細胞 (あるいはナチュラルキラーT細胞) に分化して自己細胞に応答することもできます。通常、細胞傷害性T細胞になるものはCD8+細胞で、ウイルス感染した細胞や腫瘍細胞を認識します。ナチュラルキラーT細胞 (NKTs) については後のセクションで詳述します。ここに記したT細胞や、その他のタイプのT細胞が様々な責務やエフェクター機能を様々な方法で果たすことで有害物質を認識し、他の免疫細胞を動員して排除します。また同種の侵襲に対して長期的な免疫応答を構築します。
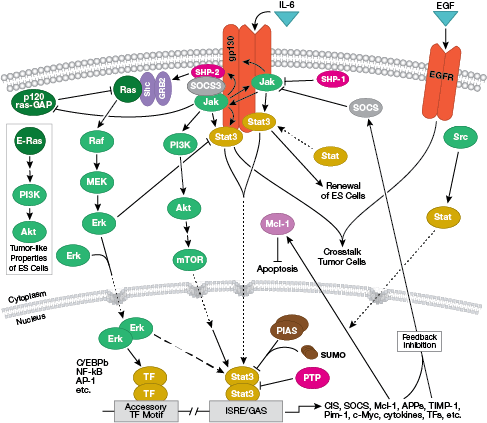
T細胞受容体シグナル伝達のインタラクティブパスウェイはこちら。
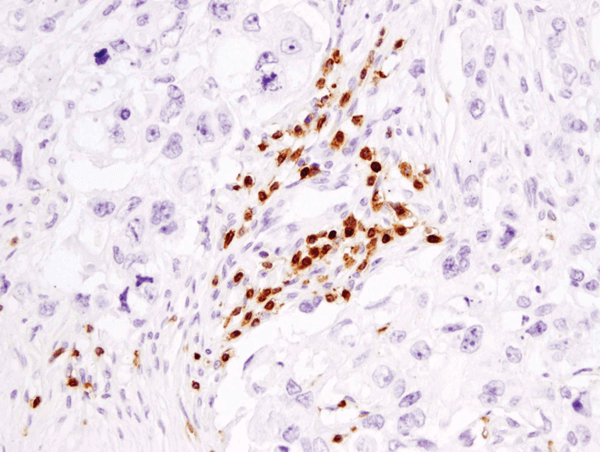
CD3ε (D7A6E) XP® Rabbit mAb #85061: パラフィン包埋ヒト乳がん組織をIHCで染色し解析しました。#85061を用いてCD3を検出することで、腫瘍組織に浸潤したT細胞を観察しました。
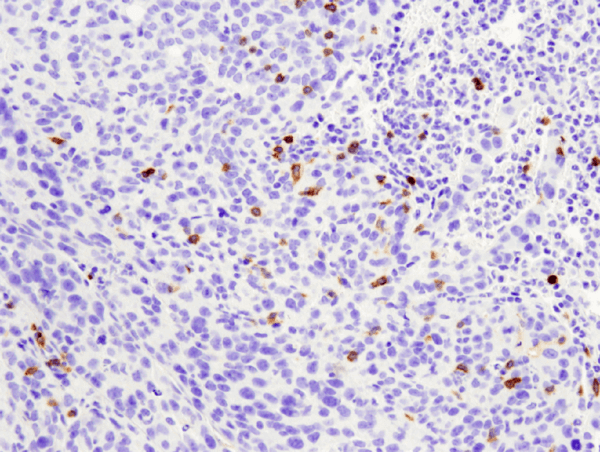
CD4 (D7D2Z) Rabbit mAb #25229: パラフィン包埋マウス4T1肺転移モデルを#25229を用いてIHCで染色し解析しました。
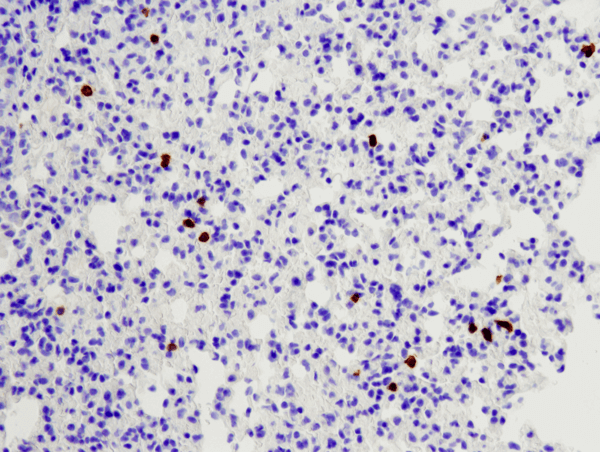
CD8α (D4W2Z) XP® Rabbit mAb (Mouse Specific) #98941: パラフィン包埋マウス肺組織を#98941を用いてIHCで染色し解析しました。
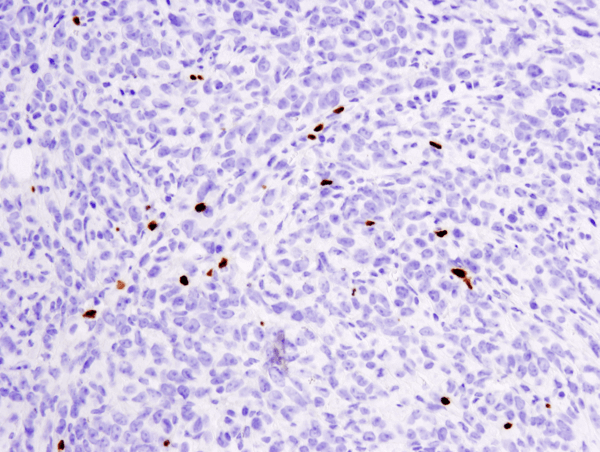
FoxP3 (D6O8R) Rabbit mAb #12653: パラフィン包埋マウス4T1肺転移モデルを#12653を用いてIHCで染色し解析しました。
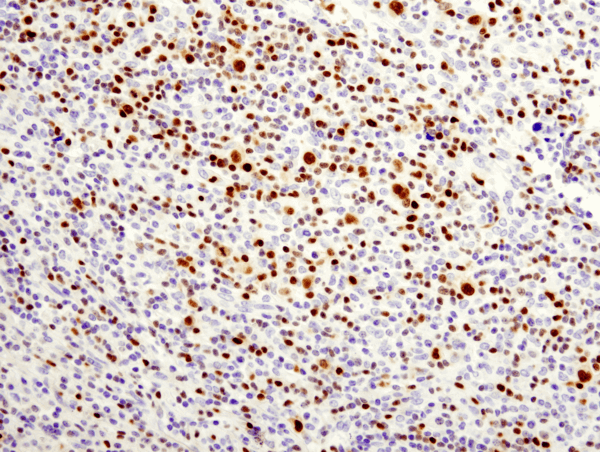
T-bet/TBX21 (D6N8B) XP® Rabbit mAb #13232: パラフィン包埋非ホジキンB細胞リンパ腫を#13232を用いてIHCで染色し解析しました。
B細胞
B細胞は血中の重要な細胞で、抗体の産生に関わっています。血中に存在する有害物質はB細胞の特異的受容体によって抗原として認識されます。抗原のプロセシング後、B細胞はT細胞の助けを得て抗体を分泌する形質細胞へと成熟します。B細胞には抗原提示やサイトカインを分泌する役割もあります。ここではB細胞の発生と機能について説明します。
末梢血にみられる白血球の15%がB細胞であるといわれています。B細胞は免疫グロブリン (Ig) を産生します。免疫グロブリンは2本の同一な重鎖と2本の同一な軽鎖からなる抗原結合タンパク質で、抗体とも呼ばれます。免疫グロブリンは特異的な抗原結合部位をもち、この部位のアミノ酸配列は抗体分子ごとに異なっています。この部位が特異性を規定し、抗体はそれぞれに異なる抗原を認識します。
上記のようにB細胞は骨髄にみられる造血幹細胞から分化します。B細胞の分化は骨髄で起こり、遺伝子の再編成を経て細胞表面にIg受容体 (抗体) が形成されます。Igを構成する重鎖と軽鎖は異なる染色体にコードされており、B細胞ではこれら別個の遺伝子がそれぞれに体細胞組換えを受けます。骨髄間質細胞に産生されるIL-7によってB細胞の分化が促進されます。発生中のB細胞では両方の染色体の重鎖と軽鎖の再編成が一定の法則に従って順番に進行し、この制御には差次的な細胞表面受容体の発現が関与しています。例えば、一方の染色体の再編成によってプレB細胞受容体が発現すると、他方の染色体の遺伝子再編成が抑制されます。これによって、成熟B細胞が単一の抗原を特異的に認識するIg受容体のみを発現するように制御されています。このプロセスは対立遺伝子排除と呼ばれます。Ig重鎖と軽鎖が適切に形成されなかった場合は、2度目の再編成による受容体編集が起こります。
細胞表面のIg受容体の発現のほか、B細胞はIg受容体の細胞外領域に抗原が結合した後に細胞内にシグナルを伝える膜貫通タンパク質も発現しています。B細胞はT細胞と同様に細胞内シグナル伝達カスケードを介して遺伝子の転写を変化させ、細胞増殖や成熟を誘導します。具体的には、まずIg受容体の細胞内ドメインがsrcファミリーキナーゼによってリン酸化されます。このシグナルはチロシンキナーゼSykとリンカータンパク質BLINKによってホスホリパーゼCとグアニンヌクレオチド交換因子に伝えられ、最終的にはプロテインキナーゼCの活性化、カルシウムの動員、Ras/Rac依存的なMAPKの活性化が起こります。
B細胞の特徴の一つにアイソタイプスイッチが挙げられます。これは具体的にはDNAの再編成によってわずかに異なる重鎖を産生させるプロセスです。また、重鎖の膜貫通領域のエクソンを選択的スプライシングで除くかどうかによって、Igを膜結合型として残すか分泌型にするかが決まります。選択的スプライシングやアイソタイプスイッチはT細胞由来のサイトカインによって誘導されます。さらに、T細胞は体細胞変異を誘導してB細胞のIgの抗原結合部位を変化させることもできます。この一見ランダムな変異によって抗原への親和性が失われた場合はB細胞の自己修正機能によって検出され、細胞死のプログラムが開始されます。したがって、T細胞とB細胞の両方を活性化させる抗原はアイソタイプスイッチと体細胞変異を介したT細胞によるB細胞の成熟を誘発し、これらがB細胞の記憶に深く関わっています。
記憶応答はそのような抗原に対する高親和性の抗体を迅速かつ大規模に産生するのが特徴です。予防接種はこの記憶応答を利用したものです。しかし、これらは自己免疫やアレルギー反応を増加させる可能性があります。
抗原によるT細胞を介さないB細胞の活性化も起こりますが、このタイプの活性化では弱い免疫記憶しか起こりません。また、この場合はB細胞を活性化するために抗原が重合体である必要があります。
注意:サイトカイン、インターロイキン (IL) は分泌性の分子で膜貫通型の細胞表面受容体に作用します。これらのサイトカインが結合すると、多くの場合Jak-STATシグナル伝達に依存した細胞内シグナル伝達パスウェイが活性化されます。
B細胞受容体シグナル伝達のインタラクティブパスウェイはこちら。
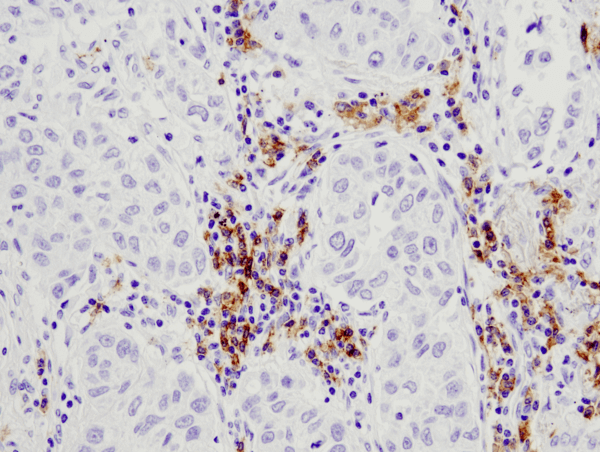
CD19 (D4V4B) XP® Rabbit mAb #90176: パラフィン包埋ヒト肺がん組織を#90176を用いてIHCで染色し解析しました。
NK細胞
ナチュラルキラー (NK) 細胞はリンパ系細胞系列に由来するもう一つのタイプの細胞です。これらはB細胞と同様に、サイトカインや骨髄間質細胞に依存して骨髄で発生します。これらは末梢血のごく一部を占めますが、抗原に特異的な受容体をもちません。代わりにNK細胞は膜貫通受容体の複雑なセットに依存しています。特にこれらはクラスI HLA (ヒトMHC) 分子を認識する抑制性の細胞表面受容体を含むため、自己のMHC分子を持つ細胞を傷害せず、クラスI HLAの発現が低下した細胞を傷害します。したがって、ウイルス感染細胞や腫瘍細胞などクラスI HLAの発現を抑制してCD8+ナチュラルキラーT (NK-T) 細胞による細胞死を回避している細胞を標的にする重要な役割を担っています。NK細胞は抗体、インターフェロン、サイトカインによっても活性化されるため、腫瘍の排除に非常に大きな役割を果たします。NK細胞は細胞質からグランザイムとパーフォリンを含む微小顆粒を放出することで作用します。これらが放出されるとパーフォリンが標的細胞表面に孔を形成し、グランザイムが標的細胞に侵入して細胞内タンパク質を分解してアポトーシスを誘導します。したがってNK細胞は細胞傷害性であると言えます。
NK-T細胞はT細胞の特殊なサブタイプで、通常はNK細胞が発現するある種の受容体やマーカーと、T細胞受容体の一部を同時に発現しています。したがって、この特殊なサブタイプはT細胞とNK細胞の性質を併せ持っています。骨髄で発生するNK細胞とは異なり、NK-T細胞は胸腺で発生します。
骨髄性細胞
骨髄系共通前駆細胞は造血幹細胞に由来する細胞で、マクロファージ、樹状細胞、その他自然免疫細胞や血液細胞など、いくつかの異なるタイプの骨髄性細胞に分化します。
マクロファージ
マクロファージは損傷した細胞や外来物質を認識して活性化され、これらを排除します。マクロファージはほとんどの組織に存在し、必要に応じて感染や死細胞に対処します。マクロファージは食作用によって認識した物質を排除します。これがこの細胞の名前の由来です (ギリシャ語で「大食らい」の意味)。マクロファージは局在する部位によって様々な形をとり、貪食以外の追加機能を果たします。
組織の損傷や感染が起こると、血中の単球が患部に動員されて分化し、マクロファージとなります。組織局在性によって異なるタイプのマクロファージがあります。例えば肝臓のクッパー細胞、肺の肺胞マクロファージ、脳のミクログリアなどです。これらの異なるタイプのマクロファージは全て単球由来ですが、常在する組織によって機能が特殊化します。一般的な貪食のほとんどは、組織常在型のマクロファージに実行されます。死細胞や異物の貪食のほか、マクロファージはサイトカインを介して他の免疫細胞にシグナルを伝えます。マクロファージはある程度、抗原提示の機能を持ち、これによってT細胞と協調して適応免疫をサポートします。また、マクロファージはIL-12などのサイトカインを分泌して局所免疫反応に重要な役割を果たします。一方で他のマクロファージはIL-10を大量に産生し、組織修復に関与しています。このように「大食らい」は貪食という主な仕事に加え、免疫系の様々な機能を担っています。
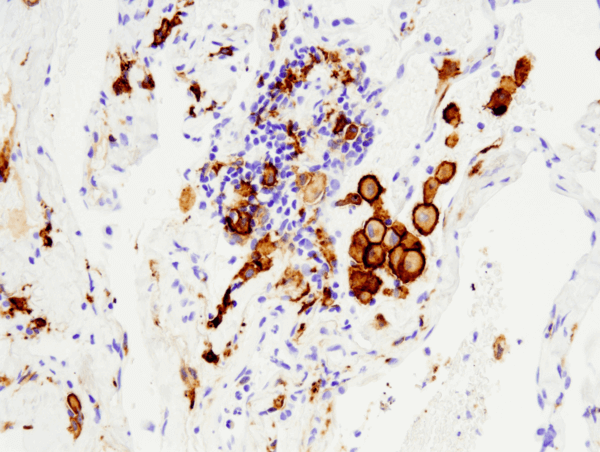
樹状細胞
樹状細胞はマクロファージと同様に、細胞や外来物質を貪食する機能がありますが、これらを消化して排除するのではなく、プロセシングを行ってT細胞に抗原として提示します。このように樹状細胞は抗原提示細胞あるいはアクセサリー細胞として機能し、自然免疫系と適応免疫系の間で病原体の情報を中継します。有害物質に手を伸ばして消化する独特の細胞突起から樹状細胞の名前が付けられました。
骨髄の造血前駆細胞から分化したばかりの未成熟な樹状細胞は、局所領域の異物をサンプリングする「学習」段階を経て、パターン認識受容体 (最も一般的なものはToll様受容体) を介して認識応答を開始します。これらの未成熟樹状細胞は皮膚組織をはじめ、外界に触れる鼻腔、胃、肺などに存在します。抗原を認識すると樹状細胞はリンパ節に移動し、そこでT細胞を訓練してその抗原を認識して適切に応答できるようにします。樹状細胞はメモリーT細胞とナイーブT細胞の両方を活性化する唯一の抗原提示細胞です。Toll様受容体は、マクロファージや樹状細胞に存在する研究の進んだ受容体のクラスで、外来病原体の認識と応答を媒介します。樹状細胞はT細胞と直接相互作用するだけでなく、IL-12などのサイトカインを介してシグナルを伝達することもできます。IL-12は免疫系に働きかけ、有害物質を攻撃する準備をします。別種の樹状細胞はサイトカインを分泌してマクロファージを動員し、過剰な廃棄物を除去している可能性があります。主要な抗原提示細胞として、樹状細胞は自然免疫系と適応免疫系のクロストークに必要不可欠な役割を果たしています。
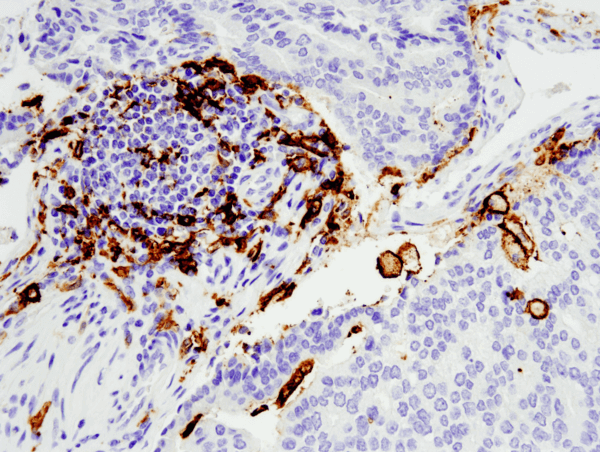
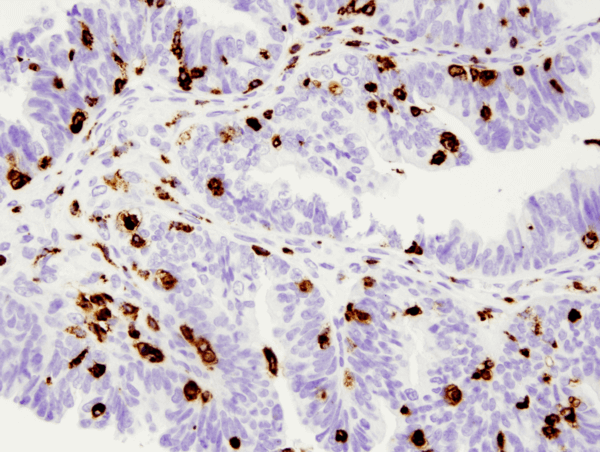
CD11c (D3V1E) XP® Rabbit mAb #45581: パラフィン包埋ヒト前立腺がん組織を#45581を用いてIHCで染色し、Leica Bond Rxプラットホームで解析しました。
その他の自然免疫細胞とシグナル伝達
その他の自然免疫系の細胞にはマスト細胞があります。この細胞は創傷治癒で病原体から保護する機能があります。病原体による刺激でマスト細胞はサイトカインやヒスタミンとヘパリンを含む顆粒を分泌します。これらはマクロファージの動員や血管の拡張など、さらなる免疫応答を媒介します。好中球、好酸球、好塩基球は細胞質に特徴的な顆粒をもち、このためまとめて顆粒球と呼ばれます。好中球の場合、この顆粒の作用で真菌や細菌などの病原体を標的とする毒性化合物の放出が起こります。
健康と疾患の免疫学
がん免疫
がんの治療に免疫細胞を利用する研究は、臨床で成功を収めたことで注目の話題となっています。最近ではPD-1など、通常T細胞の活性化や応答を抑制する、シグナルのチェックポイントタンパク質を標的とした免疫治療がフォーカスされています。また別の興味深いアプローチとして、患者から採取したT細胞に免疫編集を施す養子細胞療法が挙げられます。この方法では、遺伝子工学的操作でT細胞に患者のがん細胞に特異的なキメラ抗原受容体を発現させます。このT細胞は患者に戻すことでがん細胞を探し出し、本来のT細胞の機能に従ってこれを傷害します。これらの方法やその他免疫系を操作して有害ながん細胞を攻撃するアプローチは、画期的ながん治療法を代表するものです。