View in English?
View in English?
View in English?
CSTのシグナル伝達パスウェイ図で個々のタンパク質名をクリックすると、それに関連した研究リソースや製品情報を検索することができます。さらに、教育・研究のために、パスウェイ図をダウンロードすることも可能です。
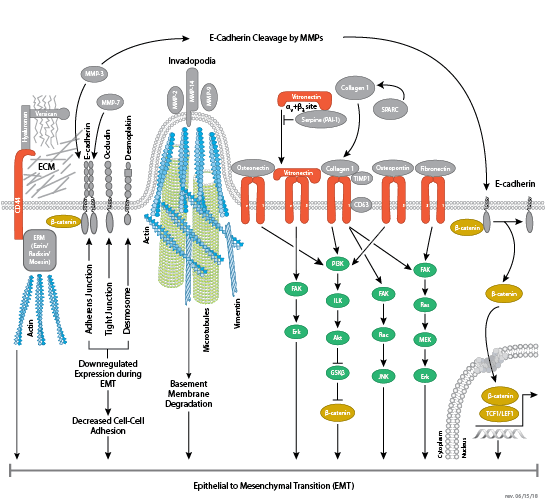
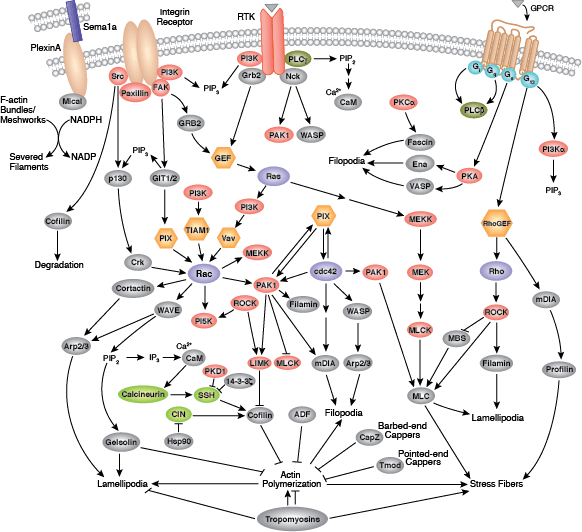
細胞は、その周辺環境の中で、細胞同士およびマトリックスとの一連の結合を形成することができます。そのような結合には、接着結合 (細胞-細胞)、密着結合 (不浸透性の細胞-細胞)、および接着斑 (細胞-マトリックス) が含まれます。
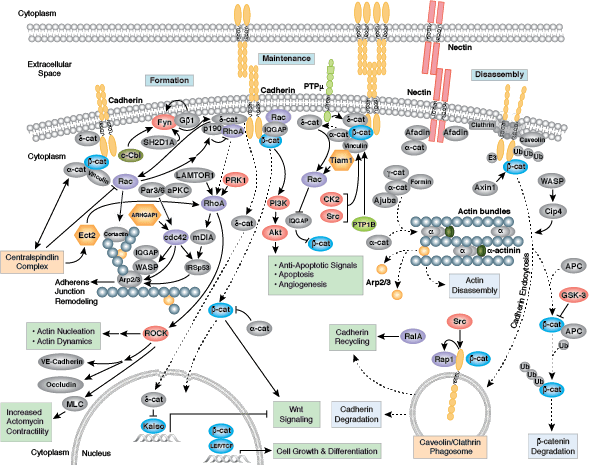
接着結合とは、結合部のタンパク質が隣接細胞の相補的な分子群と一時的に結合することで、形成、強化、伸展、退行、再形成に働く動的な構造です。接着結合の形成は、Cadherin、β-cateninおよびα-cateninから成るCadherin-Catenin複合体を介して媒介されます。細胞間結合と細胞骨格との連結は従来考えられてきた以上に動的であり、Cadherin-Catenin複合体とアクチン細胞骨格の間における複数の弱い結合や、その他の膜結合タンパク質 (Nectin、Afadinなど) に依存している可能性があります。単量体のα-cateninは接着結合部においてβ-cateninと結合しますが、遊離するとα-catenin二量体を形成して、アクチン束の形成を促進します。分岐状のアクチンネットワークから束状のアクチンフィラメントへの転換は、成熟した強い接着結合の形成、およびラメリポディアの減少と関連があります。多くの動的な細胞システムと同様に、キナーゼ、フォスファターゼ、およびアダプタータンパク質の集合がいくつかの重要なエフェクタータンパク質の活性と局在を制御します。p120 catenin (δ-catenin) は、細胞膜においてCadherinと結合して安定化します。膜結合型および細胞質型チロシンキナーゼは、弱い、もしくは新しく出来た接着部分においてβ-cateninをリン酸化します。その一方で、フォスファターゼは、成熟した接着部位においてβ-cateninとδ-cateninから付加されているリン酸を除去します。RhoファミリーGTPaseは、Cateninや他の重要な接着タンパク質の可用性や活性化状態を調節します。こうした構造タンパク質や酵素、アダプタータンパク質の集合によって、形態形成過程の一時的な結合に必要とされる、動的な細胞-細胞間接着が形成され、発生後の複雑な組織や構造の完全性が維持されます。
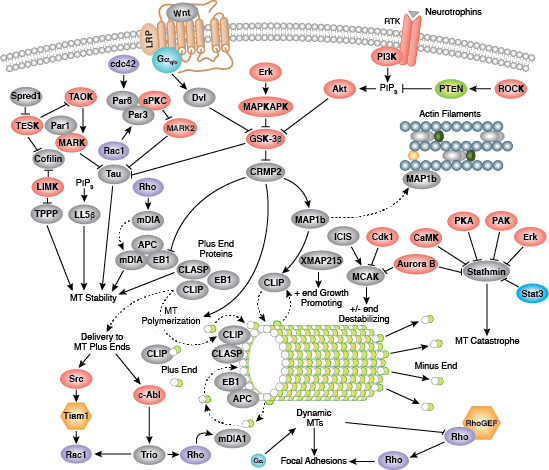
密着結合は不浸透性の細胞間接着で、上皮と内皮における液体の通過を防ぐ連続したバリアを形成します。この結合は、細胞間の透過性の制御、および細胞極性の維持、細胞頂端部と基底膜間の膜貫通タンパク質の移動の制限などの機能を担っています。密着結合は、ClaudinおよびOccludin膜貫通タンパク質から構成され、これらは細胞骨格との接合部に結合します。Occludinは、密着結合の組み立てと維持において重要であると考えられています。様々なアミノ酸残基におけるOccludinのリン酸化によって、ZO-1などの他の密着結合タンパク質との相互作用が制御されている可能性があります。
接着斑は、細胞と細胞外マトリクス (ECM) 間で形成される、主にインテグリンを介した結合です。細胞表面のα/βヘテロ二量体受容体であるインテグリンは、細胞接着および移動において重要な役割を担っています。インテグリンファミリーには少なくとも18種類のαサブユニットと8種類のβサブユニットがあり、それらは24種類の既知のインテグリンを形成し、それらのインテグリンはそれぞれ異なる組織分布を示しますが、リガンドへの特異性は重複しています。インテグリンの細胞内ドメイン尾部は、接着斑キナーゼ (FAK) など多数のシグナル伝達分子と相互作用します。インテグリンのクラスター化によってFAKが活性化することで、SrcファミリーキナーゼPI3KおよびPLCγの結合部位であるTyr397で、自己リン酸化が起こります。
View in English?